Centrifugation Can Separate Many Types of Organelles
In Chapter 3, we considered the principles of centrifugation and the uses of centrifugation techniques for separating proteins and nucleic acids. Similar approaches are used for separating and purifying various organelles, which differ in both size and density and thus undergo sedimentation at different rates.
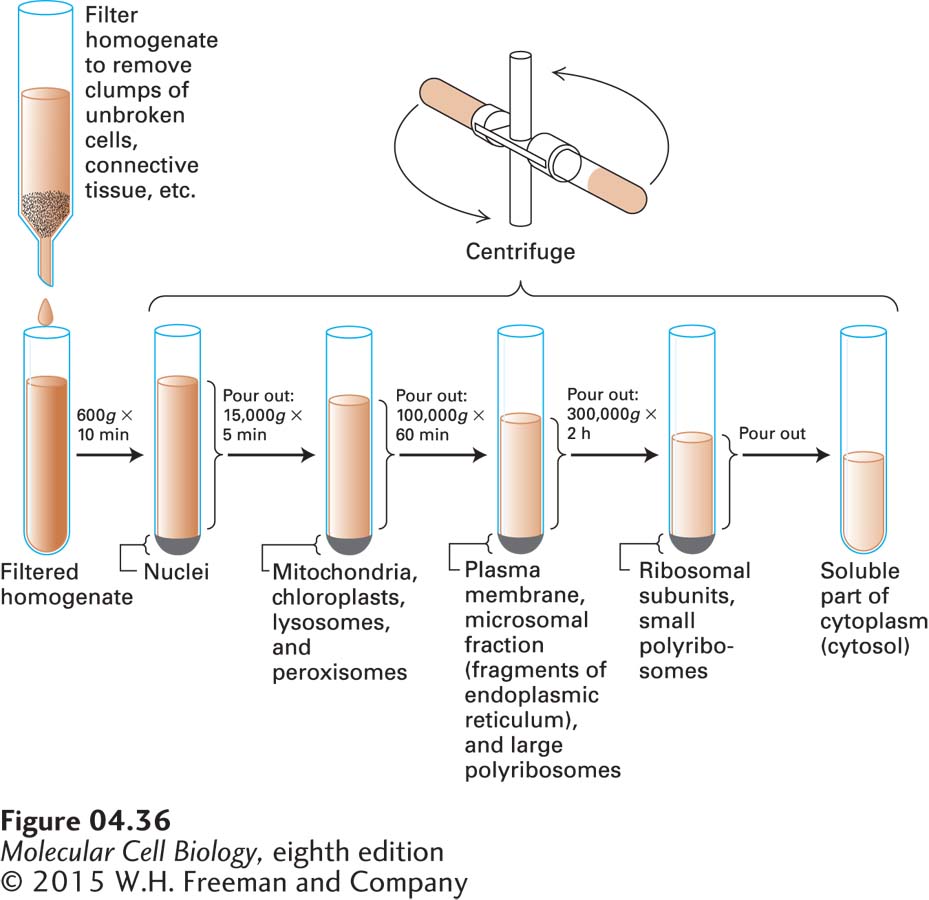
Most cell-fractionation procedures begin with differential centrifugation of a filtered cell homogenate at increasingly higher speeds (Figure 4-36). After centrifugation at each speed for an appropriate time, the liquid that remains at the top of the vessel, called the supernatant, is poured off and centrifuged at higher speed. The pelleted fractions obtained by differential centrifugation generally contain a mixture of organelles, although nuclei and viral particles can sometimes be purified completely by this procedure.
FIGURE 4-36Differential centrifugation is a common first step in fractionating a cell homogenate. The homogenate that results from disrupting cells is usually filtered to remove unbroken cells and then centrifuged at a fairly low speed to selectively pellet the nuclei—the largest organelle. The undeposited material (the supernatant) is next centrifuged at a higher speed to sediment the mitochondria, chloroplasts, lysosomes, and peroxisomes. Subsequent centrifugation in an ultracentrifuge at 100,000g for 60 minutes results in deposition of the plasma membrane, fragments of the endoplasmic reticulum, and large polyribosomes. The recovery of ribosomal subunits, small polyribosomes, and particles such as complexes of enzymes requires additional centrifugation at still higher speeds. Only the cytosol—the soluble aqueous part of the cytoplasm—remains in the supernatant after centrifugation at 300,000g for 2 hours.
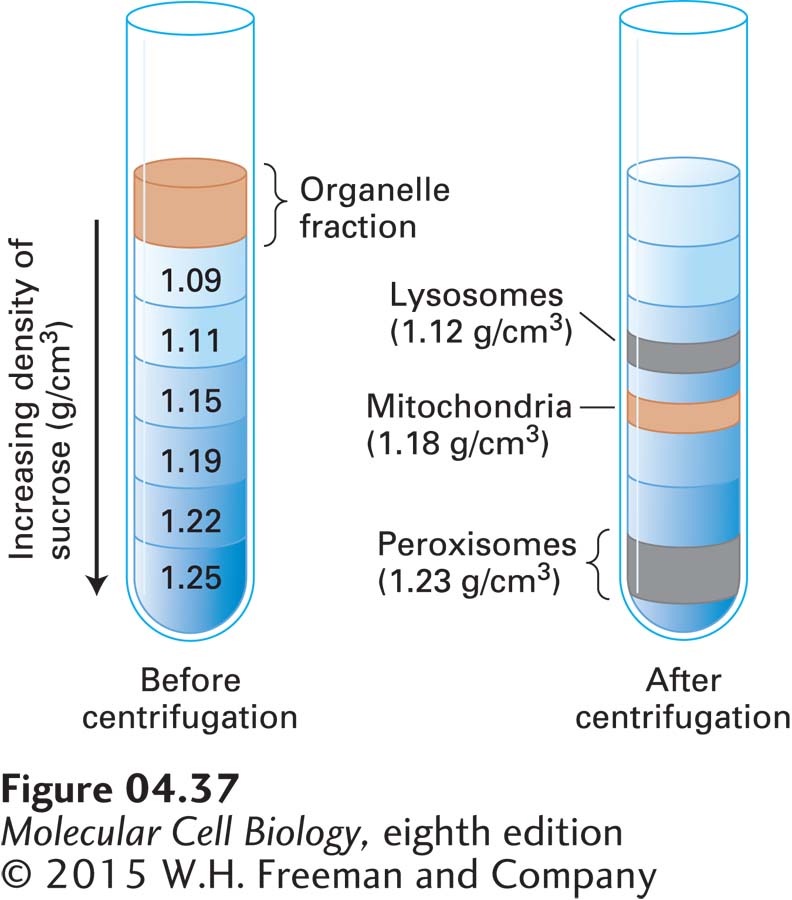
FIGURE 4-37A mixed-organelle fraction can be further separated by equilibrium density-gradient centrifugation. In this example, using rat liver, material in the pellet from centrifugation at 15,000g (see Figure 4-36) is resuspended and layered on a gradient of increasingly dense sucrose solutions in a centrifuge tube. During centrifugation for several hours, each organelle migrates to its appropriate equilibrium density and remains there. To obtain a good separation of lysosomes from mitochondria, the liver is perfused with a solution containing a small amount of detergent before the tissue is disrupted. During this perfusion period, detergent is taken into the cells by endocytosis and transferred to the lysosomes, making them less dense than they would normally be and permitting a “clean” separation of lysosomes from mitochondria.
An impure organelle fraction obtained by differential centrifugation can be further purified by equilibrium density-gradient centrifugation, which separates cellular components according to their density. After the fraction is resuspended, it is layered on top of a solution that contains a gradient of a dense non-ionic substance (e.g., sucrose or glycerol). The tube is centrifuged at a high speed (about 40,000 rpm) for several hours, allowing each particle to migrate to an equilibrium position where the density of the surrounding liquid is equal to the density of the particle (Figure 4-37). The different layers of the gradient are then recovered by pumping out the contents of the centrifuge tube through a narrow piece of tubing and collecting the fractions (see Classic Experiment 4-1).
Because each organelle has unique morphological features, the purity of organelle preparations can be assessed by examination in an electron microscope. Alternatively, organelle-specific marker molecules can be quantified. For example, the protein cytochrome c is present only in mitochondria, so the presence of this protein in a fraction of lysosomes would indicate its contamination by mitochondria. Similarly, catalase is present only in peroxisomes; acid phosphatase, only in lysosomes; and ribosomes, only in the rough endoplasmic reticulum or the cytosol.