If the many components that participate in translating mRNA had to interact in free solution, the likelihood of their coming together would be so low that the rate of amino acid polymerization would be very slow. The efficiency of translation is greatly increased by the binding of mRNA and the individual aminoacyl-tRNAs within a ribosome. The ribosome, the most abundant RNA-protein complex in the cell, directs elongation of polypeptides at a rate of 3 to 5 amino acids added per second. Small proteins of 100–200 amino acids are therefore made in a minute or less. On the other hand, it takes 2–3 hours to make the largest known protein, titin, which is found in muscle and contains about 30,000 amino acid residues. The cellular machine that accomplishes this task must be precise and persistent.
Page 189
With the aid of the electron microscope, ribosomes were first discovered as small, discrete, RNA-rich particles in cell types that secrete large amounts of protein. However, their role in protein synthesis was not recognized until reasonably pure ribosome preparations were obtained. In vitro radiolabeling experiments with such preparations showed that radioactive amino acids were incorporated into growing polypeptide chains associated with ribosomes before appearing in finished chains.
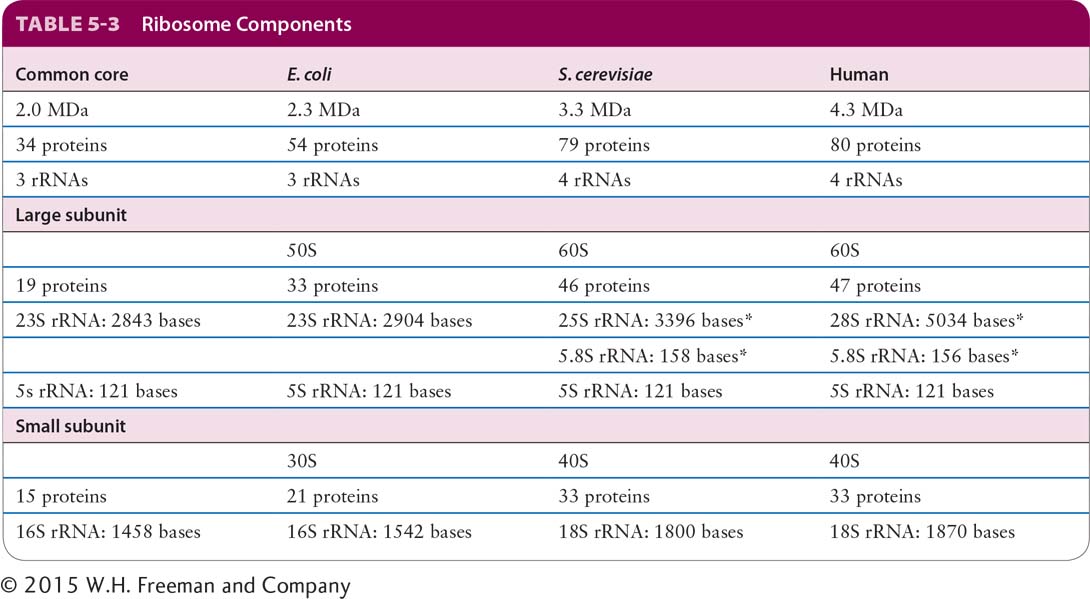
Although there are differences between the ribosomes of bacteria, archaea, and eukaryotes, the great structural and functional similarities between ribosomes from all species reflect the common evolutionary origin of the most basic constituents of living cells. A ribosome is composed of three (in bacteria and archaea) or four (in eukaryotes) different rRNA molecules and as many as 80 proteins, organized into a large subunit and a small subunit (Figure 5-22 and Table 5-3). The ribosomal subunits and the rRNA molecules are commonly designated in svedberg units (S), a measure of the sedimentation rate of macromolecules centrifuged under standard conditions—essentially, a logarithmic measure of size. The small ribosomal subunit contains a single rRNA molecule, referred to as small rRNA. The large subunit contains a molecule of large rRNA and one molecule of 5S rRNA, plus an additional molecule of 5.8S rRNA in vertebrates. The lengths of the rRNA molecules, the numbers of proteins in each subunit, and consequently, the sizes of the subunits differ between bacterial and eukaryotic cells (see Table 5-3). The assembled ribosome is 70S in bacteria and 80S in vertebrates.
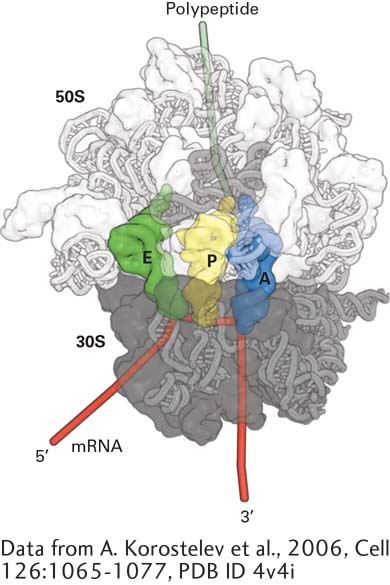
[Data from A. Korostelev et al., 2006, Cell126:1065-1077, PDB ID 4v4i.]
FIGURe 5-22Structure of the bacterial ribosome. Model of the T. thermophilus ribosome viewed along the interface between the large (50S) and small (30S) subunits. The 16S rRNA and proteins in the small subunit are dark gray. RNA is depicted as a tube model and protein surfaces are shown; the 23S rRNA and proteins in the large subunit are light gray; and the 5S rRNA is an intermediate shade of gray. The surface of the ribosome is made partially transparent to display the positions of tRNAs in the A, P, and E sites. Note that the ribosomal proteins are located primarily on the surface of the ribosome.
[Data from A. Korostelev et al., 2006, Cell126:1065-1077, PDB ID 4v4i.]
SOURCE: Data from G. Yusupov and M. Yusupov, Ann. Rev. Biochem., 2014, 83:467.
*5.8S rRNA in eukaryotes is base-paired to 25S or 28S rRNA.
SOURCE: Data from G. Yusupov and M. Yusupov, Ann. Rev. Biochem., 2014, 83:467.
Page 190
The sequences of the small and large rRNAs from several thousand organisms are now known. Although the primary nucleotide sequences of these rRNAs vary considerably, the same parts of each type of rRNA theoretically can form base-paired hairpins, stem-loops, and loop-loop interactions (see Figure 5-9), which would generate a similar three-dimensional core structure in the rRNAs and proteins of all organisms. The three-dimensional structures of bacterial and yeast ribosomes (see Figure 5-23) and of the large subunit of an archaeal ribosome have been determined by x-ray crystallography. The structures of human (see Figure 5-23) and plant ribosomes have also been determined by cryoelectron microscopy. The structure of the rRNAs in the common core, where mRNAs and tRNAs are bound and where peptide bond formation is catalyzed, is similar in all three domains of life. However, archaeal rRNAs and proteins are more similar to those of eukaryotic ribosomes than to those of bacterial ribosomes, reflecting their later divergence from a common ancestor (see Figure 1-1). For the most part, the multiple ribosomal proteins are much smaller than the rRNAs and associate with the surface of the ribosomes. Although the number of protein molecules in ribosomes greatly exceeds the number of RNA molecules, RNA constitutes about 60 percent of the mass of a bacterial ribosome, and about 50 percent of the mass of a human ribosome. Eukaryotic ribosomes are generally similar to bacterial ribosomes, but are larger because of eukaryote-specific insertions of RNA segments into regions of the common core rRNAs as well as the presence of a larger number of proteins (see Figure 5-23 and Table 5-3). Basic aspects of protein synthesis are thought to be similar among all three domains, although initiation of translation in eukaryotes, discussed later, is more complex and subject to additional mechanisms of regulation.
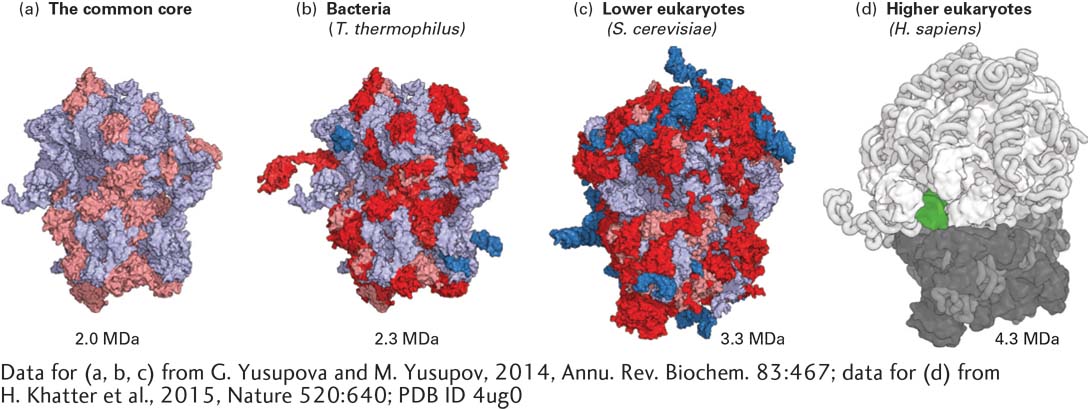
[Data for (a, b, c) from G. Yusupova and M. Yusupov, 2014, Annu. Rev. Biochem.83:467; data for (d) from H. Khatter et al., 2015, Nature520:640; PDB ID 4ug0.]
FIGURE 5-23Comparison of the common core structure at the center of ribosomes from all domains of life and bacterial, yeast, and human ribosomes. (a) RNA in the common core structure is shown in light blue and protein domains common to all ribosomes are shown in pink. Additions to the common core structure are shown in dark blue for RNA and red for proteins in ribosomes from T. thermophilus (b) and S. cerevisiae (c). Human ribosome structure (d) from cryoelectron microscopy. A tRNA visible in the E site is shown in green.
[Data for (a, b, c) from G. Yusupova and M. Yusupov, 2014, Annu. Rev. Biochem.83:467; data for (d) from H. Khatter et al., 2015, Nature520:640; PDB ID 4ug0.]
The high-resolution structures of ribosomes are providing new insights into the mechanism by which many antibiotics inhibit bacterial protein synthesis without affecting the function of mammalian ribosomes. These insights are providing important clues for the design and synthesis of new antibiotics. Such research is desperately needed as the occurrence of bacteria resistant to currently available antibiotics becomes increasingly more common, especially in hospitals, where antibiotic-resistant bacteria are under positive selection.