Eukaryotic Translation Initiation Usually Occurs at the First AUG Downstream from the 5′ End of an mRNA
During the first stage of translation, the small and large ribosomal subunits assemble around an mRNA that has a Met-tRNAiMet correctly positioned at the start codon in the ribosomal P site. In eukaryotes, the assembly of this complex is mediated by a special set of proteins known as eukaryotic translation initiation factors (eIFs). As each individual component joins the complex, it is guided by interactions with specific eIFs. Several of the initiation factors bind GTP, and the hydrolysis of GTP to GDP functions as a proofreading switch that allows subsequent steps to proceed only if the preceding step has occurred correctly. Before GTP hydrolysis, the complex is unstable, allowing dissociation of the components and a second attempt at complex formation until the correct complex assembles, resulting in GTP hydrolysis and stabilization of the appropriate complex.
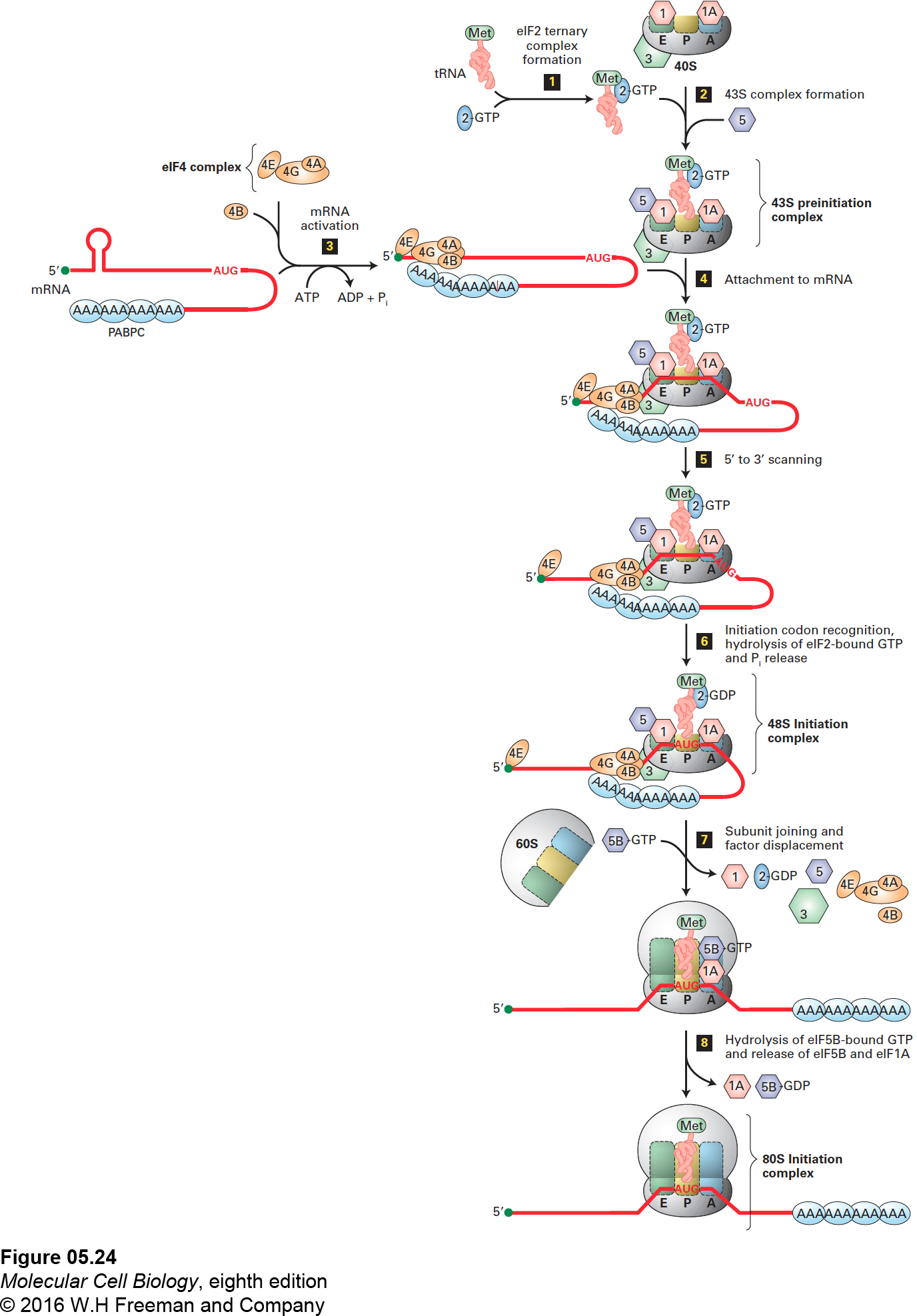
Considerable progress has been made in the past few years in understanding translation initiation in vertebrates. The current model for initiation of translation in vertebrates is depicted in Figure 5-24. Large and small ribosomal subunits released from a previous round of translation are kept apart by the binding of eIFs 1, 1A, and 3 to the small 40S subunit (Figure 5-24, top). The first step of translation initiation is formation of a 43S preinitiation complex. This preinitiation complex is formed when the 40S subunit with eIFs 1, 1A, and 3 associates with eIF5 and a ternary (three-part) complex consisting of the Met-tRNAiMet and eIF2 bound to GTP (Figure 5-24, steps 1 and 2). The initiation factor eIF2 alternates between association with GTP and GDP; it can bind Met-tRNAiMet only when it is associated with GTP. Cells can inhibit protein synthesis by phosphorylating a serine residue on the eIF2 bound to GDP; the phosphorylated complex is unable to exchange the bound GDP for GTP and cannot bind Met-tRNAiMet so protein synthesis cannot occur.
FIGURE 5-24Initiation of translation in eukaryotes. The current model of eukaryotic initiation involves eight steps. Step 1: An eIF2 ternary complex forms when eIF2·GTP binds a tRNAiMet. Step 2: When a ribosome dissociates at the termination of translation, the 40S subunit is bound by eIF1, eIF1A, and eIF3. A 43S preinitiation complex forms when this subunit associates with an eIF2 ternary complex and eIF5. Step 3: An mRNA is activated when a multisubunit eIF4 complex binds: subunit eIF4E binds to the 5′ cap, and subunit eIF4G binds multiple copies of the cytoplasmic poly(A)-binding protein (PABPC) bound to the mRNA poly(A) tail. For simplicity, binding of only one PABPC to eIF4G is shown. Then eIF4B, which stimulates eIF4A helicase activity, also joins this circular complex in which both the mRNA 5′ cap and poly(A) tail are associated with the eIF4 complex. Step 4: The 43S preinitiation complex binds an eIF4-mRNA complex. Step 5: The RNA helicase activity of subunit eIF4A unwinds any RNA secondary structure at the 5′ end of the mRNA as the 40S subunit scans in the 5′→3′ direction until it recognizes the initiation codon. For simplicity, eIF4E is diagrammed as releasing from the remainder of the eIF4 complex, but in reality, it remains associated, forming a loop in the mRNA between the 5′ cap and the scanning eIF4 complex. Step 6: Recognition of the initiation codon causes eIF5 to stimulate hydrolysis of eIF2-bound GTP. This switches the conformation of the scanning complex to a 48S initiation complex with the anticodon of tRNAiMet base-paired to the initiator AUG in the 40S-subunit P site. Step 7: The 60S subunit joins the 40S subunit, leading to the release of most of the earlier-acting eIFs as eIF5B-GTP binds to eIF1A in the ribosomal A site. The released eIF4 complex and eIF4B associate with the 5′ cap and PABPC as shown in step 3 to prepare for interaction with another 43S preinitiation complex. For simplicity, this association is not shown. Step 8: Correct association of the 40S and 60S subunits results in hydrolysis of eIF5B-bound GTP, release of eIF5B-GDP and eIF1A, and formation of the 80S initiation complex with tRNAiMet base-paired to the initiation codon in the ribosomal P site. See R. J. Jackson et al., 2010, Nature Rev. Mol. Cell Biol.11:113.
The mRNA to be translated is bound by the multisubunit eIF4 complex, which interacts with both the 5′ cap and the cytoplasmic poly(A)-binding protein (PABPC) bound in multiple copies to the mRNA poly(A) tail. Both interactions are required for translation of most mRNAs. This binding results in the formation of a circular complex (Figure 5-24, step 3). The eIF4 cap-binding complex consists of several subunits with different functions. The eIF4E subunit binds the 5′ cap on mRNAs (see Figure 5-14). The large eIF4G subunit binds cooperatively to several PABPC proteins bound to the mRNA poly(A) tail, and also forms a scaffold to which the other eIF4 subunits bind. The mRNA-eIF4 complex then associates with the preinitiation complex through an interaction between eIF4G and eIF3 (step 4).
The initiation complex then slides along, or scans, the associated mRNA as the helicase activity of eIF4A, stimulated by eIF4B, uses energy from ATP hydrolysis to unwind the RNA secondary structure (step 5). Scanning stops when the tRNAiMet anticodon recognizes the start codon, which is the first AUG downstream from the 5′ end in most eukaryotic mRNAs. Recognition of the start codon leads to hydrolysis of the GTP associated with eIF2, an irreversible step that prevents further scanning, resulting in formation of the 48S initiation complex (step 6). This commitment to the correct initiation codon is facilitated by eIF5, an eIF2 GTPase–activating protein (GAP, see Figure 3-34). Selection of the initiating AUG is facilitated by specific surrounding nucleotides called the Kozak sequence, for Marilyn Kozak, who defined it: (5′)ACCAUGG(3′). The A preceding the AUG (in bold) and the G immediately following it are the most important nucleotides affecting translation initiation efficiency.
Association of the large (60S) subunit with the small subunit, which is mediated by eIF5B bound to GTP, results in displacement of many of the initiation factors (step 7). Correct association between the ribosomal subunits results in hydrolysis of the eIF5B-bound GTP to GDP and the release of eIF5B-GDP and eIF1A (step 8), completing the formation of an 80S initiation complex. Coupling of the ribosome subunit–joining reaction to GTP hydrolysis by eIF5B allows the initiation process to continue only when the subunit interaction has occurred correctly. It also makes this an irreversible step, so that the ribosomal subunits do not dissociate until the entire mRNA is translated and protein synthesis is terminated.
The eukaryotic protein-synthesizing machinery begins translation of most cellular mRNAs within about 100 nucleotides of the 5′-capped end as just described. However, some cellular mRNAs contain an internal ribosome entry site (IRES) located far downstream from the 5′ end. It is thought that cellular IRESs form RNA structures that interact with a complex of eIF4A and eIF4G, which then associates with eIF3 bound to a 40S subunit with eIF1 and eIF1A. This assembly then binds an eIF2 ternary complex to assemble an initiation complex directly on a neighboring AUG codon. In addition, translation of some positive-stranded viral RNAs, which lack a 5′ cap, is initiated at viral IRES sequences. These RNAs fall into different classes depending on how many of the standard eIFs are required for initiation. In the case of cricket paralysis virus, the 200-nt-long IRES folds into a complex structure that interacts directly with the 40S ribosomal subunit and leads to initiation without any of the eIFs or even the initiator Met-tRNAiMet !
Page 192
In bacteria, binding of the small ribosomal subunit to an initiation site occurs by a different mechanism that allows initiation at internal sites in the polycistronic mRNAs transcribed from operons. In bacterial mRNAs, a ∼6-bp sequence complementary to the 3′ end of the small rRNA precedes the AUG start codon by 4–7 nucleotides. Base pairing between this sequence in the mRNA, called the Shine-Dalgarno sequence after its discoverers, and the small rRNA places the small ribosomal subunit in the proper position for initiation. Initiation factors comparable to eIF1A, eIF2, eIF3, and f-Met-tRNAiMet then associate with the small subunit, followed by association of the large subunit to form the complete bacterial ribosome.