During Chain Elongation Each Incoming Aminoacyl-tRNA Moves Through Three Ribosomal Sites
The correctly positioned ribosome–Met-tRNAiMet complex is now ready to begin the task of stepwise addition of amino acids by in-frame translation of the mRNA. As is the case with initiation, a set of specialized proteins, termed translation elongation factors (EFs), is required to carry out this process of chain elongation. The key steps in elongation are the entry of each succeeding aminoacyl-tRNA with an anticodon complementary to the next codon, the formation of a peptide bond, and the movement, or translocation, of the ribosome one codon at a time along the mRNA.
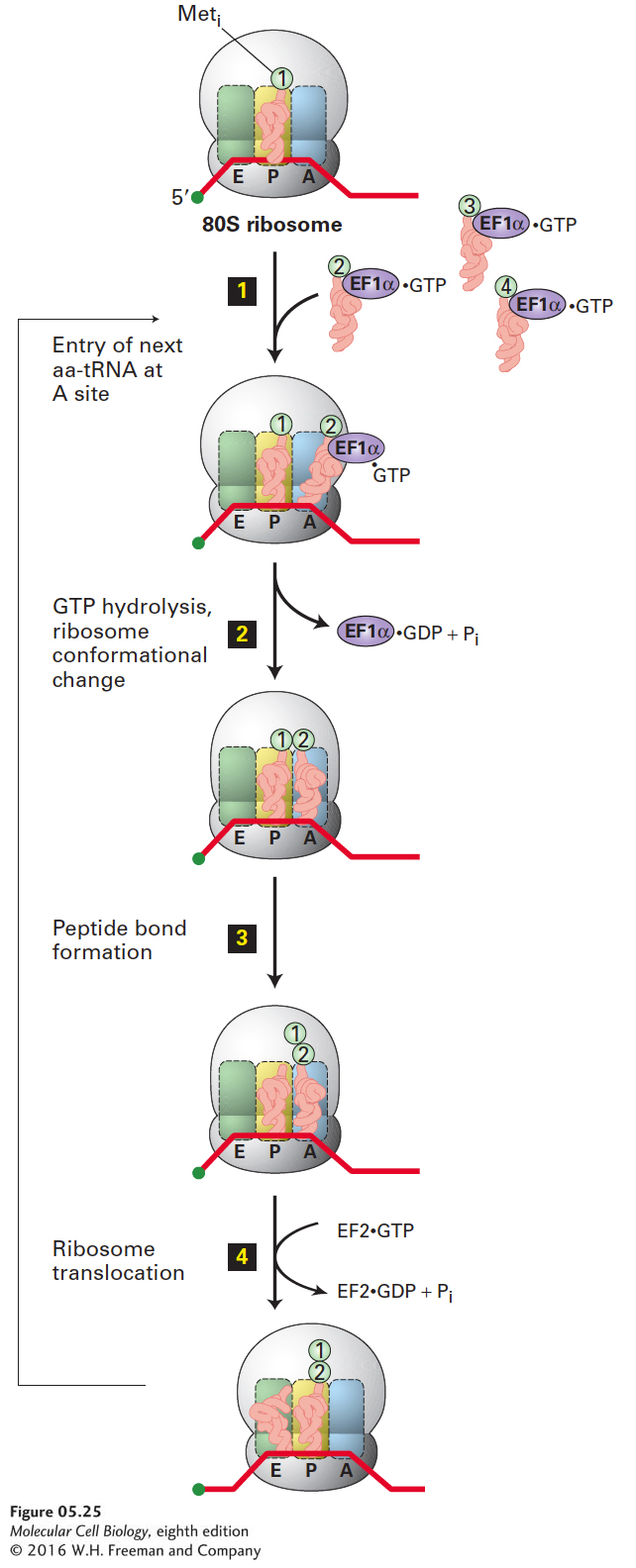
At the completion of translation initiation, as noted already, Met-tRNAiMet is bound to the P site on the assembled 80S ribosome (Figure 5-25, top). This region of the ribosome is called the P site because the tRNA chemically linked to the growing polypeptide chain is located here. The second aminoacyl-tRNA is brought into the ribosome as a ternary complex in association with EF1α·GTP and becomes bound to the A site, so named because it is where aminoacyl-tRNAs bind (step 1). EF1α·GTP bound to various aminoacyl-tRNAs diffuse into the A site, but the next step in translation proceeds only when the tRNA anticodon base-pairs with the second codon in the coding region. When that occurs properly, the GTP in the associated EF1α·GTP is hydrolyzed. The hydrolysis of GTP promotes a conformational change in EF1α that leads to release of the resulting EF1α·GDP complex and tight binding of the aminoacyl-tRNA in the A site (step 2). This conformational change also positions the aminoacylated 3′ end of the tRNA in the A site close to the 3′ end of the Met-tRNAiMet in the P site. GTP hydrolysis, and hence tight binding, does not occur if the anticodon of the incoming aminoacyl-tRNA cannot base-pair with the codon at the A site. In this case, the ternary complex diffuses away, leaving an empty A site that can associate with other aminoacyl-tRNA–EF1α·GTP complexes until a correctly base-paired tRNA is bound. Thus GTP hydrolysis by EF1α is another proofreading step that allows protein synthesis to proceed only when the correct aminoacyl-tRNA is bound to the A site. This phenomenon contributes to the fidelity of protein synthesis.
FIGURE 5-25Chain elongation in eukaryotes. Once the 80S ribosome with Met-tRNAiMet in the ribosome P site is assembled (top), a ternary complex bearing the second amino acid (aa2) coded by the mRNA binds to the A site (step 1). Following a conformational change in the ribosome induced by hydrolysis of GTP in EF1α·GTP (step 2), the large rRNA catalyzes peptide bond formation between Meti and aa2 (step 3). Hydrolysis of GTP in EF2·GTP causes another conformational change in the ribosome that results in its translocation one codon along the mRNA and shifts the unacylated tRNAiMet to the E site and the tRNA with the bound peptide to the P site (step 4). The cycle can begin again with binding of a ternary complex bearing aa3 to the now open A site. In the second and subsequent elongation cycles, the tRNA at the E site is ejected during step 2 as a result of the conformational change induced by hydrolysis of GTP in EF1α·GTP.
Page 194
With the initiating Met-tRNAiMet at the P site and the second aminoacyl-tRNA tightly bound at the A site, the α-amino group of the second amino acid reacts with the “activated” (ester-linked) methionine on the initiator tRNA, forming a peptide bond (Figure 5-25, step 3; see Figure 5-17). This peptidyltransferase reaction is catalyzed by the large rRNA, which precisely orients the interacting atoms, permitting the reaction to proceed. The 2′-hydroxyl of the terminal A of the peptidyl-tRNA in the P site also participates in catalysis. The catalytic ability of the large rRNA in bacteria has been demonstrated by carefully removing the vast majority of the protein from large ribosomal subunits. The nearly pure bacterial 23S rRNA can catalyze a peptidyltransferase reaction between analogs of aminoacyl-tRNA and peptidyl-tRNA. Further support for the catalytic role of large rRNA in protein synthesis came from high-resolution crystallographic studies showing that no proteins lie near the site of peptide bond synthesis in the crystal structure of the bacterial large subunit.
Following peptide bond synthesis, the ribosome translocates a distance equal to one codon along the mRNA. This translocation step is monitored by hydrolysis of the GTP in eukaryotic EF2·GTP. Once translocation has occurred correctly, the bound GTP is hydrolyzed, another irreversible process that prevents the ribosome from moving along the RNA in the wrong direction or from translocating an incorrect number of nucleotides. As a result of conformational changes in the ribosome that accompany proper translocation and the resulting GTP hydrolysis by EF2, tRNAiMet, now without its activated methionine, is moved to the E (exit) site on the ribosome; concurrently, the second tRNA, now covalently bound to a dipeptide (a peptidyl-tRNA), is moved to the P site (Figure 5-25, step 4). Translocation thus returns the ribosome conformation to a state in which the A site is open and able to accept another aminoacyl-tRNA complexed with EF1α·GTP, beginning another cycle of chain elongation.
Repetition of the elongation cycle depicted in Figure 5-25 adds amino acids one at a time to the carboxyl terminus of the growing polypeptide as directed by the mRNA sequence until a stop codon is encountered. In subsequent cycles, the conformational change that occurs in step 2 ejects the unacylated tRNA from the E site. As the nascent polypeptide chain becomes longer, it threads through a channel in the large ribosomal subunit, exiting at a position opposite the side that interacts with the small subunit (Figures 5-22, 5-26).
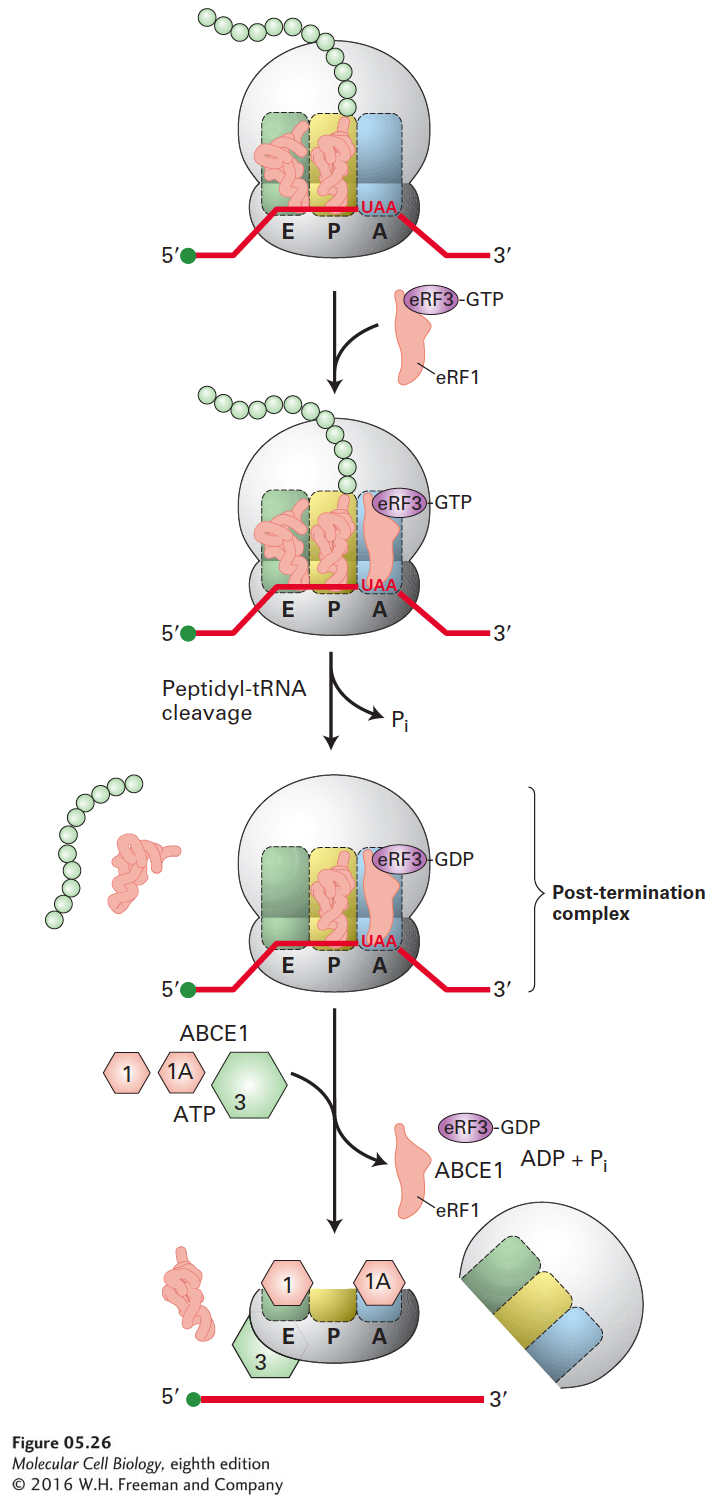
FIGURE 5-26Termination of translation in eukaryotes. When a ribosome bearing a nascent protein chain reaches a stop codon (UAA, UGA, UAG), release factor eRF1 enters the A site together with eRF3·GTP. Hydrolysis of the bound GTP is accompanied by cleavage of the peptide chain from the tRNA in the P site and ejection of the tRNA in the E site, forming a post-termination complex. The ribosomal subunits are separated by the action of the ABCE1 ATPase together with eIF1, eIF1A, and eIF3. The 40S subunit is released bound to these eIFs, ready to initiate another cycle of translation (see Figure 5-24).
In the absence of the ribosome, the three-base-pair RNA-RNA hybrid between the tRNA anticodons and the mRNA codons in the A and P sites would not be stable; RNA-RNA duplexes between separate RNA molecules must be considerably longer to be stable under physiological conditions. However, multiple interactions between the large and small rRNAs and the general domains of tRNAs (e.g., the D and TψCG loops, see Figure 5-20) stabilize the tRNAs in the A and P sites, while other RNA-RNA interactions sense correct codon-anticodon base pairing, ensuring that the genetic code is read properly. Then, interactions between rRNAs and the general domains of all tRNAs result in the movement of the tRNAs between the A, P, and E sites as the ribosome translocates along the mRNA one three-nucleotide codon at a time.