Now that we have seen how the genetic information encoded in the nucleotide sequence of DNA is translated into the proteins and RNAs that perform most cell functions, we can appreciate the necessity for precisely copying DNA sequences during DNA replication, in preparation for cell division (see Figure 5-1, step 4). The regular pairing of bases in the double-helical DNA structure suggested to Watson and Crick that new DNA strands are synthesized by using the existing (parent) strands as templates in the formation of new, daughter strands that are complementary to the parent strands.
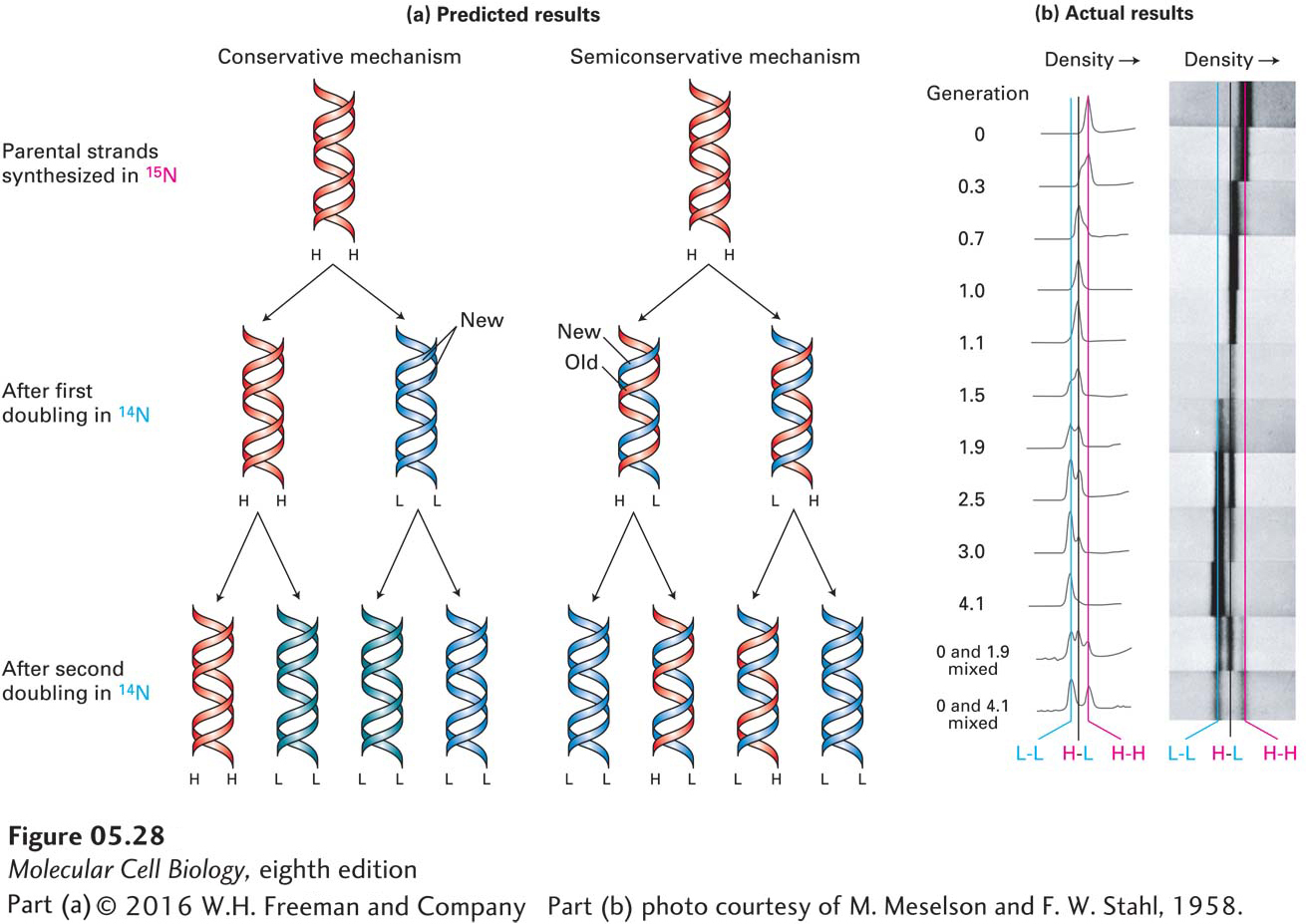
This base-pairing template model theoretically could proceed by either a conservative or a semiconservative mechanism. If a conservative mechanism were used, the two daughter strands would form a new double-stranded DNA molecule and the parent duplex would remain intact. If a semiconservative mechanism were used, the parent strands would be permanently separated and each would form a duplex molecule with the newly synthesized daughter strand base-paired to it. Definitive evidence that duplex DNA is replicated by a semiconservative mechanism came from a now classic experiment conducted by M. Meselson and W. F. Stahl, outlined in Figure 5-28.
[Part (b) photo courtesy of M. Meselson and F. W. Stahl, 1958.]
EXPERIMENTAL FIGURE 5-28The Meselson-Stahl experiment. This experiment showed that DNA replicates by a semiconservative mechanism. E. coli cells were initially grown in a medium containing ammonium salts prepared with “heavy” nitrogen (15N) until all the cellular DNA was labeled. After the cells were transferred to a medium containing the normal “light” isotope (14N), samples were removed periodically from the cultures, and the DNA in each sample was analyzed by equilibrium density-gradient centrifugation, a procedure that separates macromolecules on the basis of their density. This technique can separate heavy-heavy (H-H), light-light (L-L), and heavy-light (H-L) duplexes into distinct bands. (a) Expected composition of daughter duplex molecules synthesized from 15N-labeled DNA after cells are shifted to 14N-containing medium if DNA replication occurs by a conservative or by a semiconservative mechanism. Parent heavy (H) strands are in red; light (L) strands synthesized after shift to 14N-containing medium are in blue. Note that the conservative mechanism would never generate H-L DNA, and that the semiconservative mechanism would never generate H-H DNA but would generate H-L DNA during the first and subsequent doublings. With additional replication cycles, the 15N-labeled (H) strands from the original DNA would be diluted, so that the vast bulk of the DNA would consist of L-L duplexes with either mechanism. (b) Actual banding patterns of DNA subjected to equilibrium density-gradient centrifugation before and after 15N-labeled cells were shifted to 14N-containing medium. DNA bands were visualized under UV light and photographed. The traces on the left are a measure of the density of the photographic signal, and hence the DNA concentration, along the length of the centrifuged cells from left to right. The number of generations (far left) following the shift to 14N-containing medium was determined by counting the concentration of cells in the culture. This value corresponds to the number of DNA replication cycles that had occurred at the time each sample was taken. After one generation of growth, all the extracted DNA had the density of H-L DNA. After 1.9 generations, approximately half the DNA had the density of H-L DNA; the other half had the density of L-L DNA. With additional generations, a larger and larger fraction of the extracted DNA consisted of L-L duplexes; H-H duplexes never appeared. These results match the predicted pattern for the semiconservative replication mechanism depicted in (a). The bottom two centrifuge cells contained mixtures of H-H DNA and DNA isolated at 1.9 and 4.1 generations in order to clearly show the positions of H-H, H-L, and L-L DNA in the density gradient. See M. Meselson and F. W. Stahl, 1958, Proc. Nat’l Acad. Sci. USA 44:671.
[Part (b) photo courtesy of M. Meselson and F. W. Stahl, 1958.]
Copying of a DNA template strand into a complementary strand is thus a common feature of DNA replication, transcription of DNA into RNA, and as we will see later in this chapter, DNA repair and recombination. In all cases, the information in the template, in the form of the specific sequence of nucleotides, is preserved. In some viruses, single-stranded RNA molecules function as templates for the synthesis of complementary RNA or DNA strands. However, the vast preponderance of RNA and DNA in cells is synthesized from preexisting duplex DNA.