Two Systems Use Recombination to Repair Double-Strand Breaks in DNA
Ionizing radiation (e.g., x- and γ-radiation) and some anticancer drugs cause double-strand breaks in DNA. These lesions are particularly severe because incorrect rejoining of double strands of DNA can lead to gross chromosomal rearrangements that can affect the functioning of genes. For example, incorrect joining could create a “hybrid” gene that codes for the N-terminal portion of one amino acid sequence fused to the C-terminal portion of a completely different amino acid sequence; or a chromosomal rearrangement could bring the promoter of one gene close to the coding region of another gene, changing the level or cell type in which that gene is expressed.
Page 208
Two systems have evolved to repair double-strand breaks: homologous recombination, discussed below, and nonhomologous end joining (NHEJ), which is error-prone, since several nucleotides are invariably lost at the point of repair.
Error-Prone Repair by Nonhomologous End Joining The predominant mechanism for repairing double-strand breaks in multicellular organisms involves rejoining of the nonhomologous ends of two DNA molecules. Even if the joined DNA fragments come from the same chromosome, the repair process results in the loss of several base pairs at the joining point (Figure 5-39). The formation of such a possibly mutagenic deletion is one example of how repair of DNA damage can introduce mutations.
[Part (b) data from J. R. Walker, R. A. Corpina, and J. Goldberg, 2001, Nature412:607, PDB ID 1jey.]
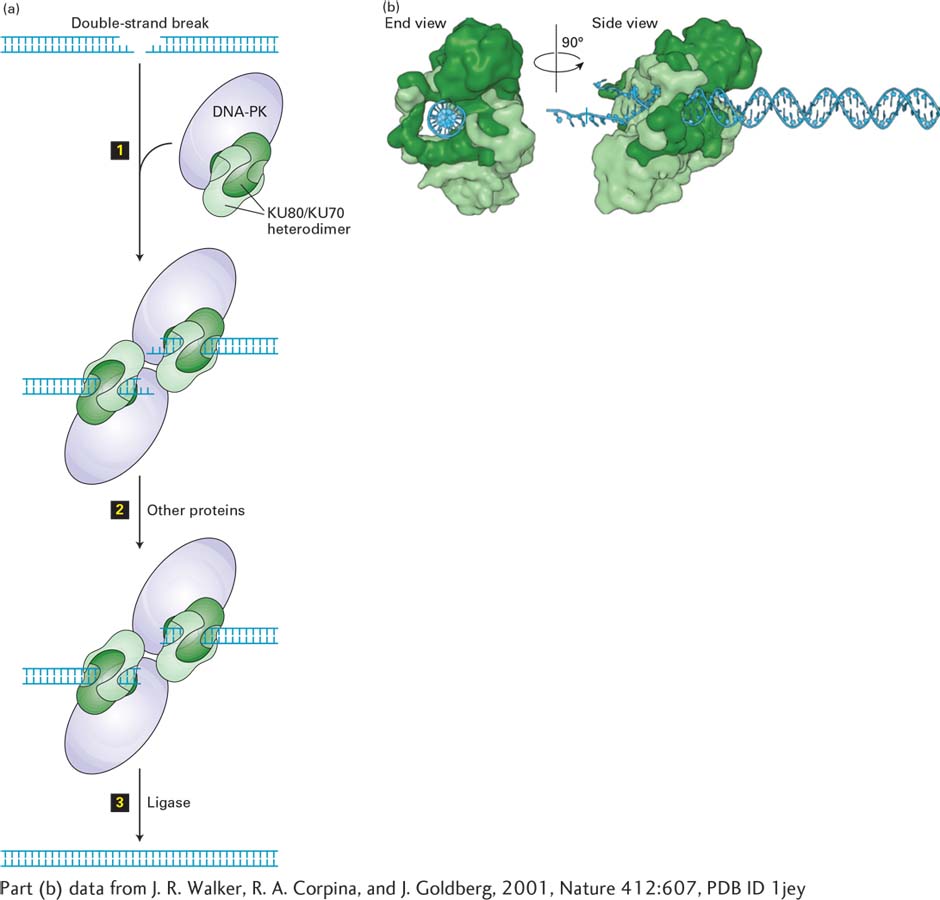
FIGURE 5-39Nonhomologous end joining. (a) When sister chromatids are not available to help repair double-strand breaks, nucleotide sequences are butted together that were not apposed in the unbroken DNA. These DNA ends are usually from the same chromosome locus, and when linked together, several base pairs are lost. Occasionally, ends from different chromosomes are accidentally joined together. A complex of two proteins, Ku and DNA-dependent protein kinase (DNA-PK), binds to the ends of a double-strand break (step 1). After formation of a synapse, the ends are further processed by nucleases, resulting in removal of a few bases (step 2), and the two double-stranded molecules are ligated together (step 3). As a result, the double-strand break is repaired, but several base pairs at the site of the break are removed. See G. Chu, 1997, J. Biol. Chem.272:24097; M. Lieber et al., 1997, Curr. Opin. Genet. Devel.7:99; and D. van Gant et al., 2001, Nature Rev. Genet. 2:196. (b) Structure of KU70/KU80 bound to a duplex DNA end. The complex is shown in a view down the DNA axis (left) and from the side (right). KU80 is light green, KU70 dark green, DNA blue.
[Part (b) data from J. R. Walker, R. A. Corpina, and J. Goldberg, 2001, Nature412:607, PDB ID 1jey.]
Since diffusion of DNA within the viscous nucleoplasm is fairly slow, the correct ends of a broken chromosome are generally rejoined together, albeit with a loss of base pairs, before they diffuse too far apart for nonhomologous end joining to be efficient. Since most of the length of most human genes consists of introns that are spliced out of the processed mRNA, in most cases the small deletion generated is not harmful. Occasionally, however, broken ends from different chromosomes are joined together, leading to translocation of pieces of DNA from one chromosome to another. Such translocations may generate chimeric genes that can have drastic effects on normal cell function, such as uncontrollable cell growth, which is the hallmark of cancer (see Figure 8-38). The devastating effects of double-strand breaks make these the “most unkindest cuts of all,” to borrow a phrase from Shakespeare’s Julius Caesar.