Segregation of Mutations in Breeding Experiments Reveals Their Dominance or Recessivity
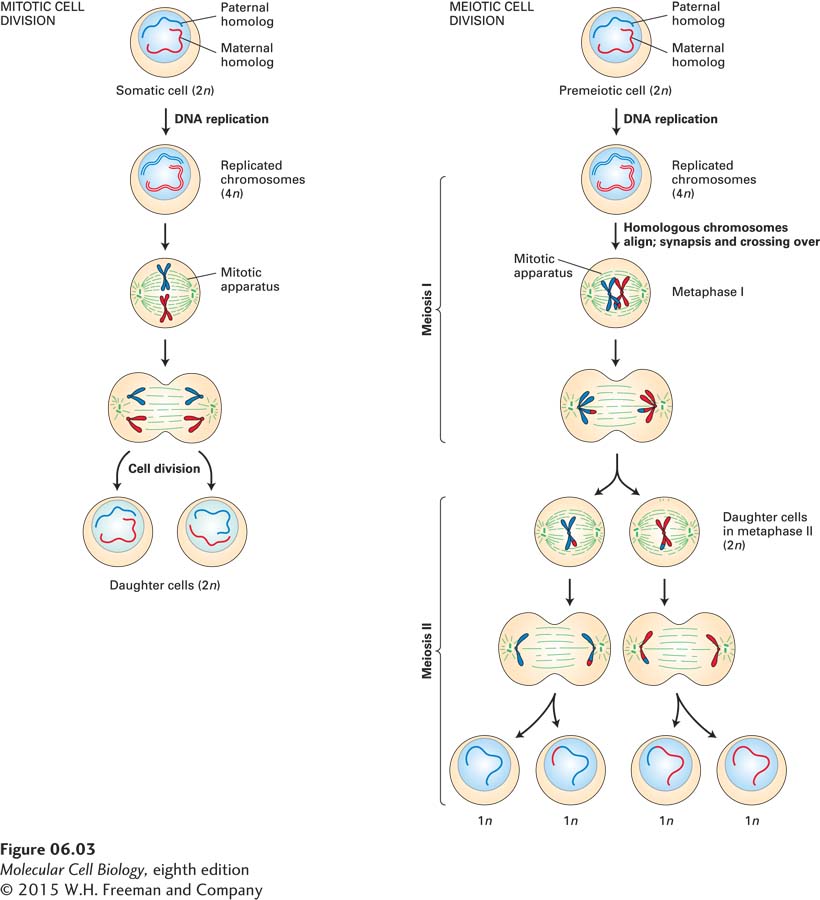
Geneticists exploit the normal life cycle of an organism to test for the dominance or recessivity of alleles. To see how this is done, we must first review the type of cell division that gives rise to gametes (sperm and egg cells in higher plants and animals). Whereas the body (somatic) cells of most multicellular organisms divide by mitosis, the germ cells that give rise to gametes undergo meiosis. Like somatic cells, pre-meiotic germ cells are diploid, containing two homologs of each morphological type of chromosome. The two homologs that constitute each pair of homologous chromosomes are descended from different parents, and thus their genes may exist in different allelic forms. Figure 6-3 depicts the major events in mitotic and meiotic cell division. In mitosis, DNA replication is always followed by cell division, yielding two diploid daughter cells. In meiosis, one round of DNA replication is followed by two separate cell divisions, yielding haploid (1n) cells known as gametes that contain only one chromosome of each homologous pair. The apportionment, or segregation, of the replicated homologous chromosomes to daughter cells during the first meiotic division is random and different chromosomes segregate independently of one another, yielding gametes with different mixes of paternal and maternal chromosomes.
Page 226
FIGURE 6-3Comparison of mitosis and meiosis. Both somatic cells and pre-meiotic germ cells have two copies of each chromosome (2n), one maternal and one paternal. In mitosis, the replicated chromosomes, each composed of two sister chromatids, align at the cell center in such a way that both daughter cells receive a maternal and a paternal homolog of each morphological type of chromosome. During the first meiotic division, however, each replicated chromosome pairs with its homologous partner at the cell center; this pairing off is referred to as synapsis, and crossing over between homologous chromosomes is evident at this stage. One replicated chromosome of each morphological type then goes into each daughter cell. The resulting cells undergo a second division without intervening DNA replication, so that one of the sister chromatids of each morphological type is apportioned to the daughter cells. In the second meiotic division, the alignment of chromatids and their equal segregation into daughter cells is the same as in mitotic division. The alignment of pairs of homologous chromosomes in metaphase I is random with respect to other chromosome pairs, resulting in a mix of paternally and maternally derived chromosomes in each daughter cell.
Page 227
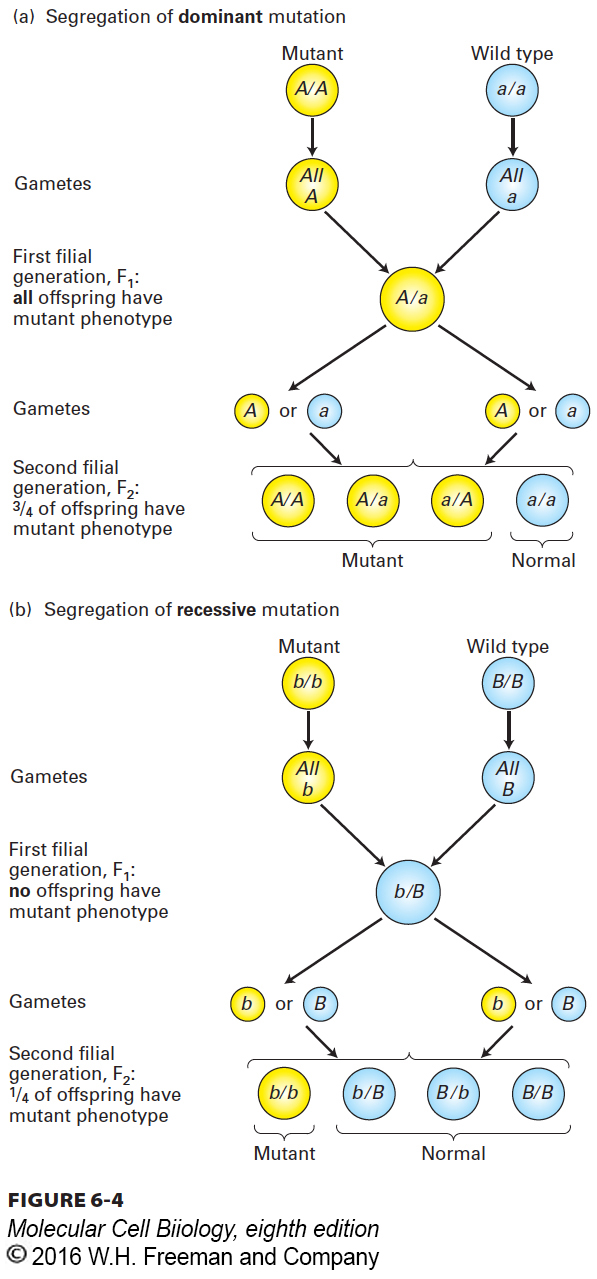
As a way to avoid unwanted complexity, geneticists usually strive to begin breeding experiments with strains of organisms that are homozygous for the genes under examination. In such true-breeding strains, every individual will receive the same allele from each parent, and therefore the composition of alleles will not change from one generation to the next. When a true-breeding mutant strain is mated to a true-breeding wild-type strain, all the first filial (F1) progeny will be heterozygous (Figure 6-4). If the F1 progeny exhibit the mutant trait, then the mutant allele is dominant; if the F1 progeny exhibit the wild-type trait, then the mutant allele is recessive. Further crossing between F1 individuals will also reveal different patterns of inheritance according to whether the mutation is dominant or recessive. When F1 individuals that are heterozygous for a dominant allele are crossed among themselves, three-fourths of the resulting F2 progeny will exhibit the mutant trait. In contrast, when F1 individuals that are heterozygous for a recessive allele are crossed among themselves, only one-fourth of the resulting F2 progeny will exhibit the mutant trait.
FIGURE 6-4Segregation patterns of dominant and recessive mutations in crosses between true-breeding strains of diploid organisms. All the offspring in the first (F1) generation are heterozygous. If the mutant allele is dominant, the F1 offspring will exhibit the mutant phenotype, as in part (a). If the mutant allele is recessive, the F1 offspring will exhibit the wild-type phenotype, as in part (b). Crossing of the F1 heterozygotes among themselves also produces different segregation ratios for dominant and recessive mutant alleles in the F2 generation.
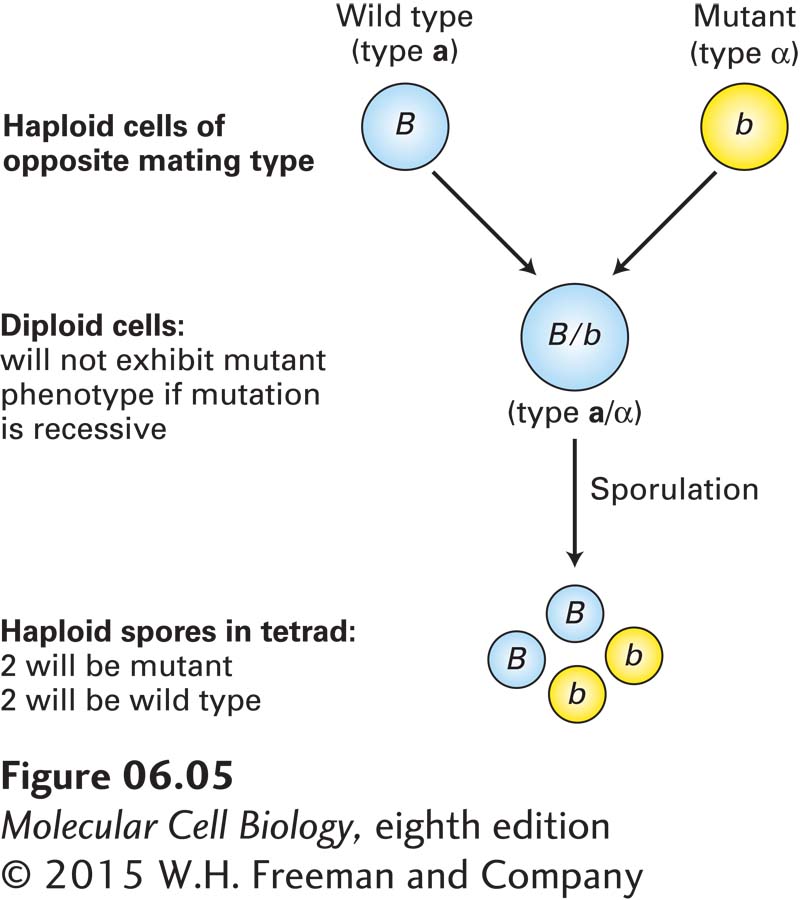
As noted earlier, the yeast S. cerevisiae, an important experimental organism, can exist in either a haploid or a diploid state. In these unicellular eukaryotes, crosses between haploid cells can determine whether a mutant allele is dominant or recessive. Haploid yeast cells, which carry one copy of each chromosome, can be of two different mating types, known as a and α. Haploid cells of opposite mating type can mate to produce a/α diploids, which carry two copies of each chromosome. If a new mutation with an observable phenotype is isolated in a haploid strain, the mutant strain can be mated to a wild-type strain of the opposite mating type to produce a/α diploids that are heterozygous for the mutant allele. If these diploids exhibit the mutant trait, then the mutant allele is dominant, but if the diploids exhibit the wild-type trait, then the mutant allele is recessive. When a/α diploids are placed under starvation conditions, the cells undergo meiosis, each giving rise to a tetrad of four haploid spores, two of type a and two of type α. Sporulation of a heterozygous diploid cell yields two spores carrying the mutant allele and two carrying the wild-type allele (Figure 6-5). Under appropriate conditions, yeast spores will germinate, producing vegetative haploid strains of both mating types.
FIGURE 6-5Segregation of alleles in yeast. Haploid Saccharomyces cells of opposite mating type (i.e., one of mating type a and one of mating type α) can mate to produce an a/α diploid. If one haploid carries a dominant wild-type allele and the other carries a recessive mutant allele of the same gene, the resulting heterozygous diploid will express the dominant trait. Under certain conditions, a diploid cell will form a tetrad of four haploid spores. Two of the spores in the tetrad will express the recessive trait and two will express the dominant trait.