Proteins Can Be Removed from Membranes by Detergents or High-Salt Solutions
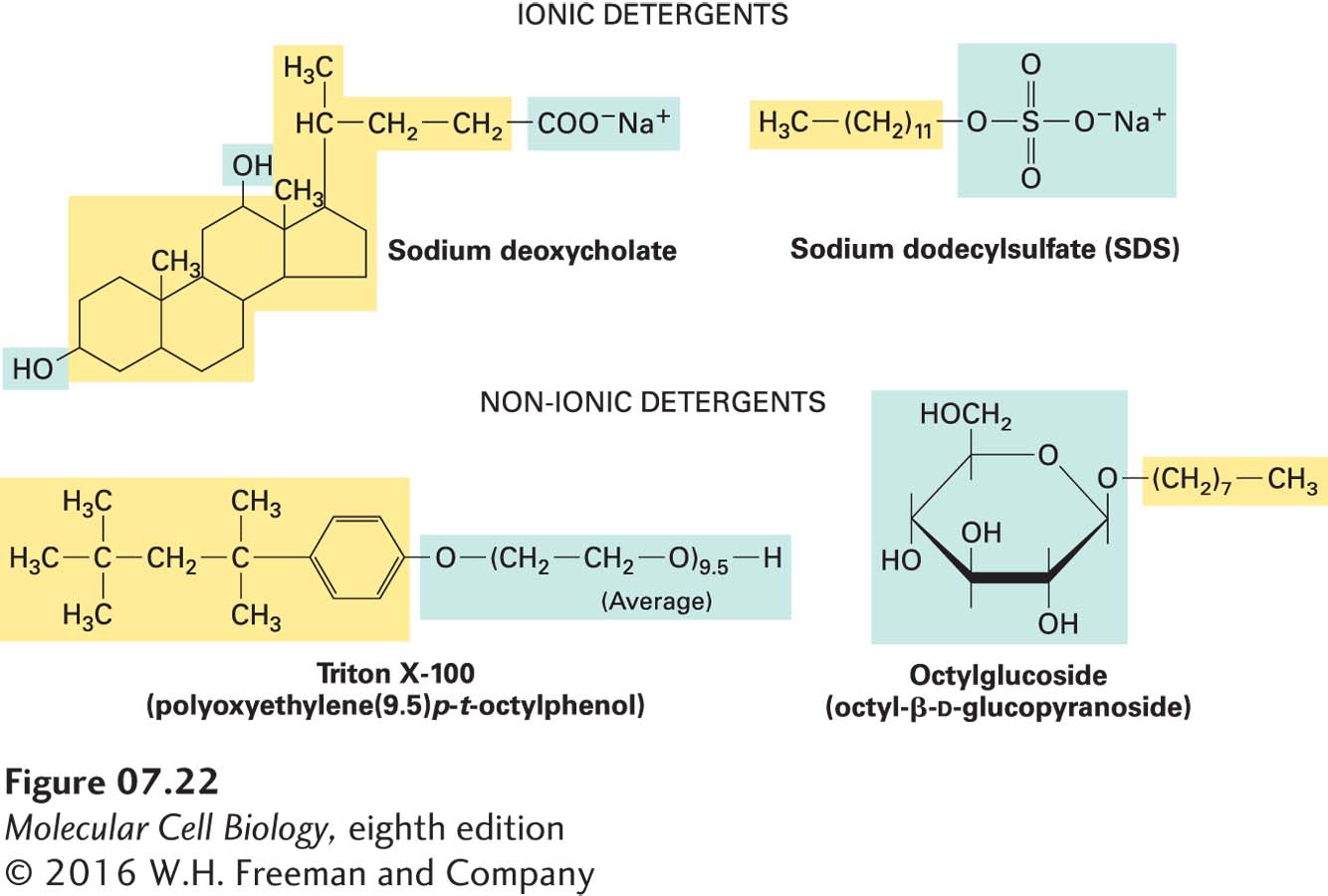
Membrane proteins are often difficult to purify and study, mostly because of their tight association with membrane lipids and other membrane proteins. Detergents, which are amphipathic molecules that disrupt membranes by intercalating into phospholipid bilayers, can be used to solubilize lipids and many membrane proteins. The hydrophobic part of a detergent molecule is attracted to the phospholipid hydrocarbons and mingles with them readily; the hydrophilic part is strongly attracted to water. Some detergents, such as bile salts, are natural products, but most are synthetic molecules developed for cleaning and for dispersing mixtures of oil and water in the food industry (e.g., creamy peanut butter) (Figure 7-22). Ionic detergents, such as sodium deoxycholate (a bile salt) and sodium dodecylsulfate (SDS), contain a charged group; non-ionic detergents, such as Triton X-100 and octylglucoside, lack a charged group. At very low concentrations, detergents dissolve in pure water as isolated molecules. As the concentration increases, the molecules begin to form micelles—small spherical aggregates in which the hydrophilic parts of the molecules face outward and the hydrophobic parts cluster in the center (see Figure 7-3c). The critical micelle concentration (CMC) at which micelles form is characteristic of each detergent and is a function of the structures of its hydrophobic and hydrophilic parts.
FIGURE 7-22Structures of four common detergents. The hydrophobic part of each molecule is shown in yellow; the hydrophilic part, in blue. The bile salt sodium deoxycholate is a natural product; the others are synthetic. Although ionic detergents commonly cause denaturation of proteins, non-ionic detergents do not and are thus useful in solubilizing integral membrane proteins.
Page 291
Ionic and non-ionic detergents interact differently with proteins and have different uses in the lab. Ionic detergents bind to the exposed hydrophobic regions of membrane proteins as well as to the hydrophobic cores of water-soluble proteins. Because of their charge, these detergents can disrupt ionic and hydrogen bonds. At high concentrations, sodium dodecylsulfate, for example, completely denatures proteins by binding to every side chain, a property that is exploited in SDS gel electrophoresis (see Figure 3-38). Non-ionic detergents generally do not denature proteins and are thus useful in extracting proteins in their folded and active form from membranes before the proteins are purified. Protein-protein interactions, especially the weaker ones, can be sensitive to both ionic and non-ionic detergents.
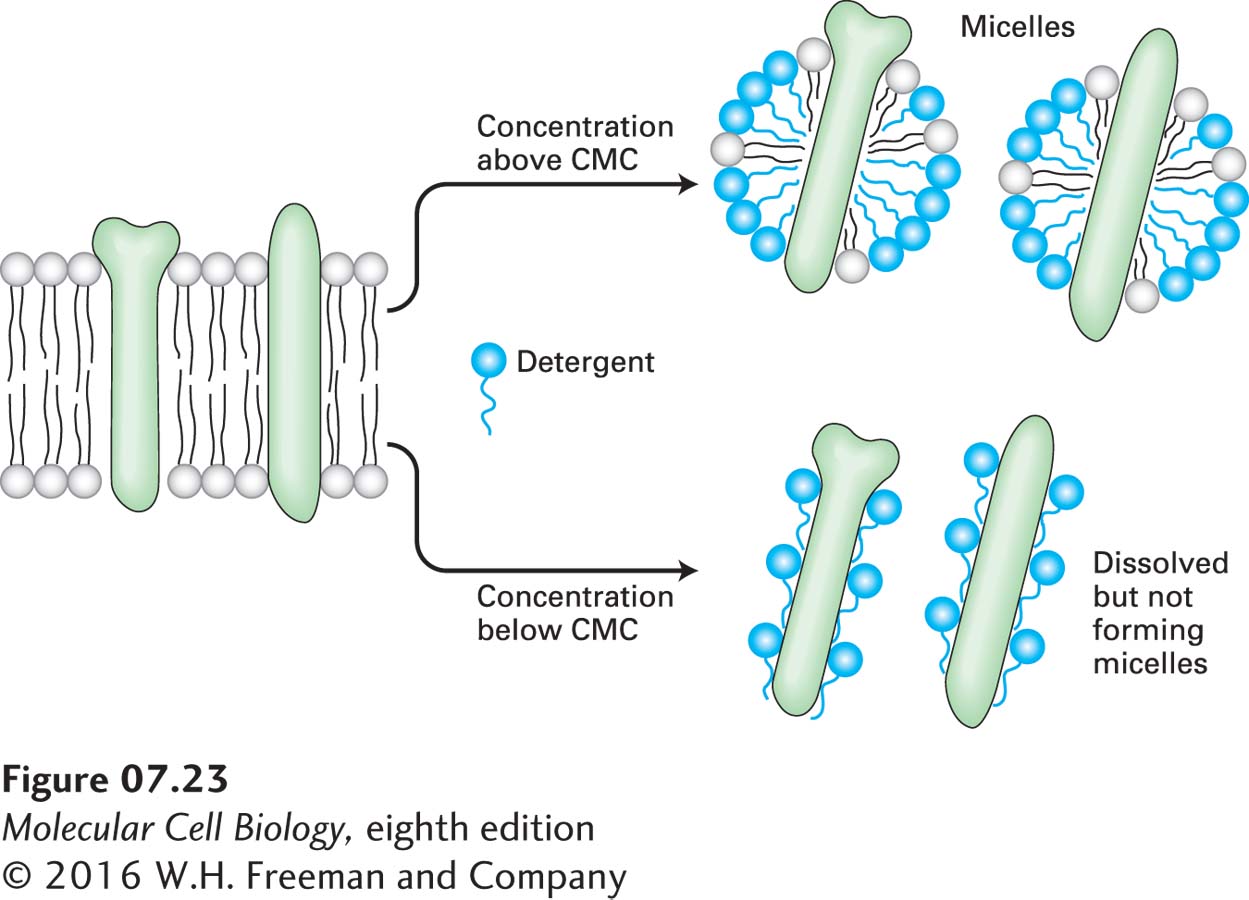
At high concentrations (above the CMC), non-ionic detergents solubilize biological membranes by forming mixed micelles of detergent, phospholipids, and integral membrane proteins, bulky hydrophobic structures that do not dissolve in water (Figure 7-23, top). At low concentrations (below the CMC), these detergents bind to the hydrophobic regions of most integral membrane proteins, but without forming micelles, allowing the proteins to remain soluble (Figure 7-23, bottom). Creating such an aqueous solution of integral membrane proteins is a necessary first step in protein purification.
FIGURE 7-23Solubilization of integral membrane proteins by non-ionic detergents. At a concentration higher than its critical micelle concentration (CMC), a detergent solubilizes lipids and integral membrane proteins, forming mixed micelles containing detergent, protein, and lipid molecules. At concentrations below the CMC, non-ionic detergents (e.g., octylglucoside, Triton X-100) can dissolve membrane proteins without forming micelles by coating the membrane-spanning regions.
Page 292
Treatment of cultured cells with a buffered salt solution containing a non-ionic detergent such as Triton X-100 extracts water-soluble proteins as well as integral membrane proteins from cellular membranes. As noted earlier, the exoplasmic and cytosolic domains of integral membrane proteins are generally hydrophilic and soluble in water. The membrane-spanning domains, however, are rich in hydrophobic and uncharged residues (see Figure 7-14). When separated from membranes, these exposed hydrophobic segments tend to interact with one another, causing the protein molecules to aggregate and precipitate from aqueous solutions. The hydrophobic parts of non-ionic detergent molecules preferentially bind to the hydrophobic segments of transmembrane proteins, preventing protein aggregation and allowing the proteins to remain in aqueous solution. Detergent-solubilized transmembrane proteins can then be purified by affinity chromatography and other techniques used in purifying water-soluble proteins (see Chapter 3).
As discussed previously, most peripheral membrane proteins are bound to specific transmembrane proteins or membrane phospholipids by ionic or other weak noncovalent interactions. Generally, peripheral proteins can be removed from the membrane by solutions of high ionic strength (high salt concentrations), which disrupt ionic bonds, or by chemicals that bind divalent cations such as Mg2+. Unlike integral membrane proteins, most peripheral proteins are soluble in water and need not be solubilized by non-ionic detergents.