DNA Transposons Are Present in Prokaryotes and Eukaryotes
Most mobile elements in bacteria transpose directly as DNA. In contrast, most mobile elements in eukaryotes are retrotransposons, but eukaryotic DNA transposons also occur. Indeed, the original mobile elements discovered by Barbara McClintock are DNA transposons.
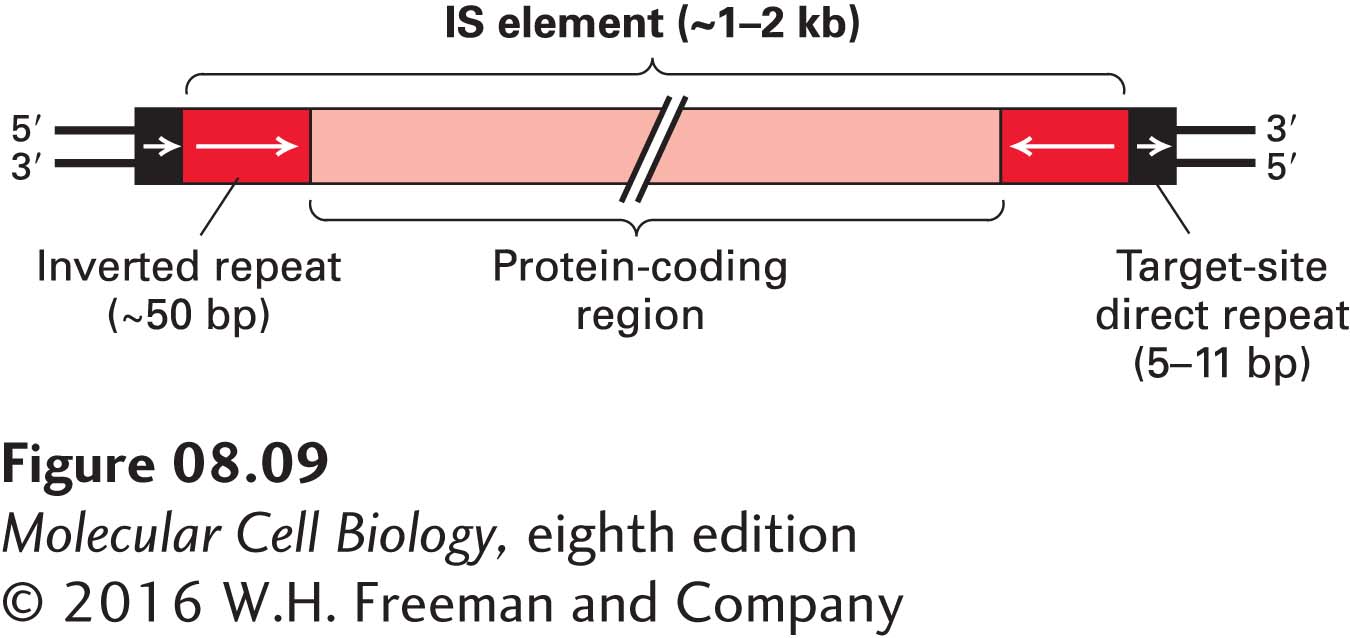
Bacterial Insertion Sequences The first molecular understanding of mobile elements came from the study of certain E. coli mutations caused by the spontaneous insertion of a DNA sequence, about 1–2 kb long, into the middle of a gene. These inserted stretches of DNA are called insertion sequences, or IS elements. So far, more than 1000 different IS elements have been found in E. coli and other bacteria.
Transposition of an IS element is a very rare event, occurring in only one in 105–107 cells per generation, depending on the particular IS element. Often transpositions inactivate essential genes, killing the host cell and the IS elements it carries. Therefore, higher rates of transposition would probably result in too great a mutation rate for the host organism to survive. However, since IS elements transpose more or less randomly, some transposed sequences enter nonessential regions of the host genome (e.g., regions between genes), allowing the host cell to survive. At a very low rate of transposition, most host cells survive and therefore propagate the symbiotic IS element. IS elements can also insert themselves into plasmids or lysogenic viruses and can thus be transferred to other cells. In this way, IS elements can transpose into the chromosomes of new host cells.
FIGURE 8-9General structure of bacterial IS elements. The relatively large central region of an IS element, which encodes one or two enzymes required for transposition, is flanked by an inverted repeat at each end. The sequences of the inverted repeats are nearly identical, but they are oriented in opposite directions. The sequence is characteristic of a particular IS element. The 5′ and 3′ short direct (as opposed to inverted) repeats are not transposed with the insertion element; rather, they are insertion-site sequences that become duplicated, with one copy at each end, during insertion of a mobile element. The length of the direct repeats is constant for a given IS element, but their sequence depends on the site of insertion and therefore varies with each transposition of the IS element. Arrows indicate sequence orientation. The regions in this diagram are not to scale; the coding region makes up most of the length of an IS element.
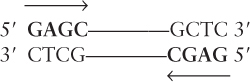
The general structure of IS elements is diagrammed in Figure 8-9. An inverted repeat of 10–40 bp is invariably present at each end of an IS element. In an inverted repeat, the 5′→3′ sequence on one strand is repeated on the other strand, such as
Between the inverted repeats is a region that encodes a transposase, an enzyme required for transposition of the IS element to a new site. The transposase is expressed very rarely, accounting for the very low frequency of transposition. An important hallmark of IS elements is the presence of a short direct repeat sequence, containing 5–11 bp, depending on the particular IS element, immediately adjacent to both ends of the inserted element. The length of the direct repeat is characteristic of each type of IS element, but its sequence depends on the target site where a particular copy of the IS element inserted. When the sequence of a mutated gene containing an IS element is compared with the wild-type gene sequence, only one copy of the direct repeat is found in the wild-type gene. Duplication of this target-site sequence to create the second direct repeat adjacent to an IS element occurs during the insertion process.
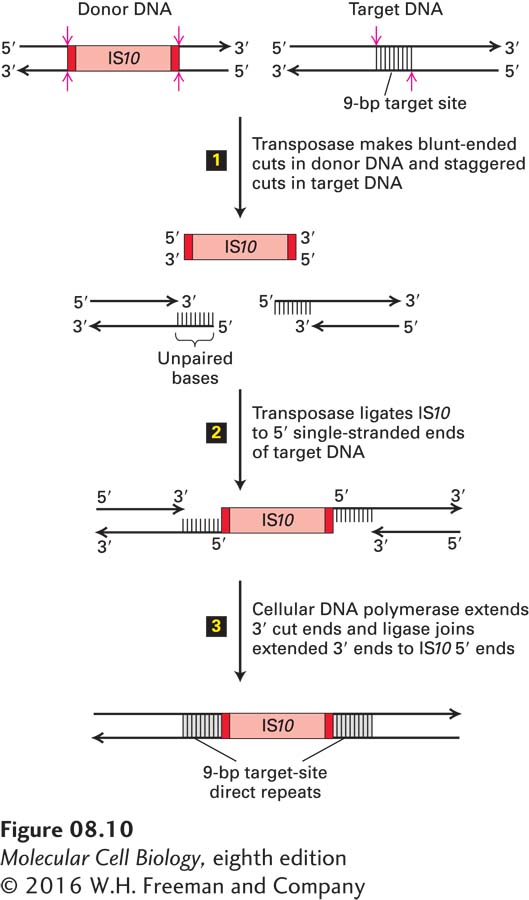
As depicted in Figure 8-10, transposition of an IS element occurs by a “cut-and-paste” mechanism. Transposase performs three functions in this process: it (1) precisely excises the IS element from the donor DNA, (2) makes staggered cuts in a short sequence in the target DNA, and (3) ligates the 3′ termini of the IS element to the 5′ ends of the cut donor DNA. Finally, a host-cell DNA polymerase fills in the single-stranded gaps, generating the short direct repeats that flank IS elements, and DNA ligase joins the free ends.
FIGURE 8-10Model for transposition of bacterial insertion sequences. Step 1: Transposase, which is encoded by the IS element (IS10 in this example), cleaves both strands of the donor DNA next to the inverted repeats (dark red), excising the IS10 element. At a largely random target site, transposase makes staggered cuts in the target DNA. In the case of IS10, the two cuts are 9 bp apart. Step 2: Ligation of the 3′ ends of the excised IS element to the staggered sites in the target DNA is also catalyzed by transposase. Step 3: The 9-bp gaps of single-stranded DNA left in the resulting intermediate are filled in by a cellular DNA polymerase; finally, cellular DNA ligase forms the 3′→5′ phosphodiester bonds between the 3′ ends of the extended target DNA strands and the 5′ ends of the IS10 strands. This process results in duplication of the target-site sequence on each side of the inserted IS element. Note that the lengths of the target site and IS10 are not to scale. See H. W. Benjamin and N. Kleckner, 1989, Cell59:373; and 1992, Proc. Natl. Acad. Sci. USA89:4648.
Eukaryotic DNA Transposons McClintock’s original discovery of mobile elements came from her observation of spontaneous mutations in maize that affect the production of enzymes required to make anthocyanin, a purple pigment in maize kernels. Mutant kernels are white and wild-type kernels are purple. One class of these mutations is revertible at high frequency, whereas a second class does not revert unless they occur in the presence of the first class of mutations. McClintock called the agents responsible for the first class of mutations activator (Ac) elements and those responsible for the second class dissociation (Ds) elements because they also tended to be associated with chromosome breaks.
Many years after McClintock’s pioneering discoveries, cloning and sequencing revealed that Ac elements are equivalent to bacterial IS elements. Like IS elements, they contain inverted terminal repeat sequences that flank a region encoding a transposase, which recognizes the terminal repeats and catalyzes transposition to a new site in the host DNA. Ds elements are deleted forms of Ac elements in which a portion of the sequence encoding transposase is missing. Because it does not encode a functional transposase, a Ds element cannot move by itself. However, in plants that carry Ac elements and thus express a functional transposase, Ds elements can be transposed because they retain the inverted terminal repeats recognized by the transposase.
Page 315
Since McClintock’s early work on mobile elements in corn, DNA transposons have been identified in other eukaryotes. For instance, approximately half of all the spontaneous mutations observed in Drosophila are due to the insertion of mobile elements. Although most of the mobile elements in Drosophila function as retrotransposons, at least one—the P element—functions as a DNA transposon, moving by a mechanism similar to that used by bacterial insertion sequences. Current methods for constructing transgenic Drosophila depend on engineered, high-level expression of the P-element transposase and use of the P-element inverted terminal repeats as targets for transposition, as discussed in Chapter 6 (see Figure 6-20).
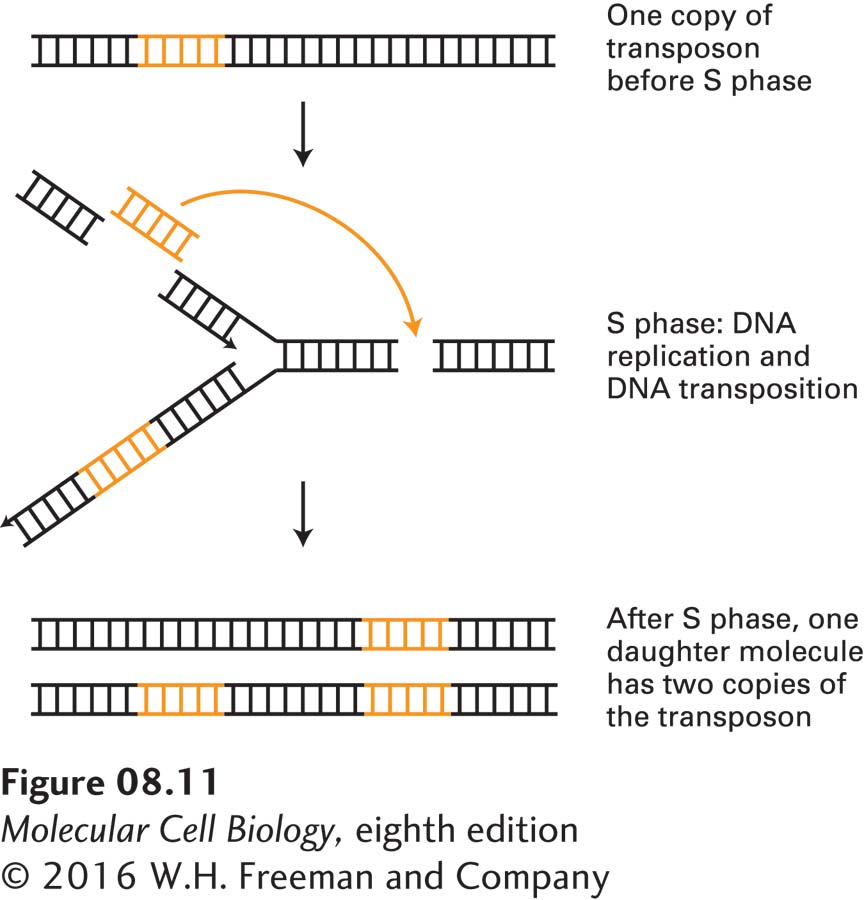
DNA transposition by the cut-and-paste mechanism can result in an increase in the copy number of a transposon if it occurs during the S phase of the cell cycle (see Figure 1-21), when DNA synthesis occurs. Such an increase happens when the donor DNA is in one of the two daughter DNA molecules in a region of a chromosome that has replicated but the target DNA is in a region that has not yet replicated. When DNA replication is complete at the end of the S phase, the target DNA in its new location has also been replicated, resulting in a net increase in the total number of transposon copies in the cell (Figure 8-11). When such a transposition occurs during the S phase preceding meiosis, one of the four germ cells produced contains the extra copy of the transposon. Repetition of this process over evolutionary time has resulted in the accumulation of large numbers of DNA transposons in the genomes of some organisms. Human DNA contains about 300,000 copies of full-length and deleted DNA transposons, amounting to about 3 percent of human DNA. As we will see shortly, this mechanism can lead to the transposition of genomic DNA as well as the transposon itself.
FIGURE 8-11Mechanism for increasing DNA-transposon copy number. If a DNA transposon, which transposes by a cut-and-paste mechanism (see Figure 8-10), transposes during S phase from a region of the chromosome that has replicated to a region that has not yet replicated, then when chromosomal replication is completed, one of the two daughter chromosomes will have a net increase of one copy of the transposon.