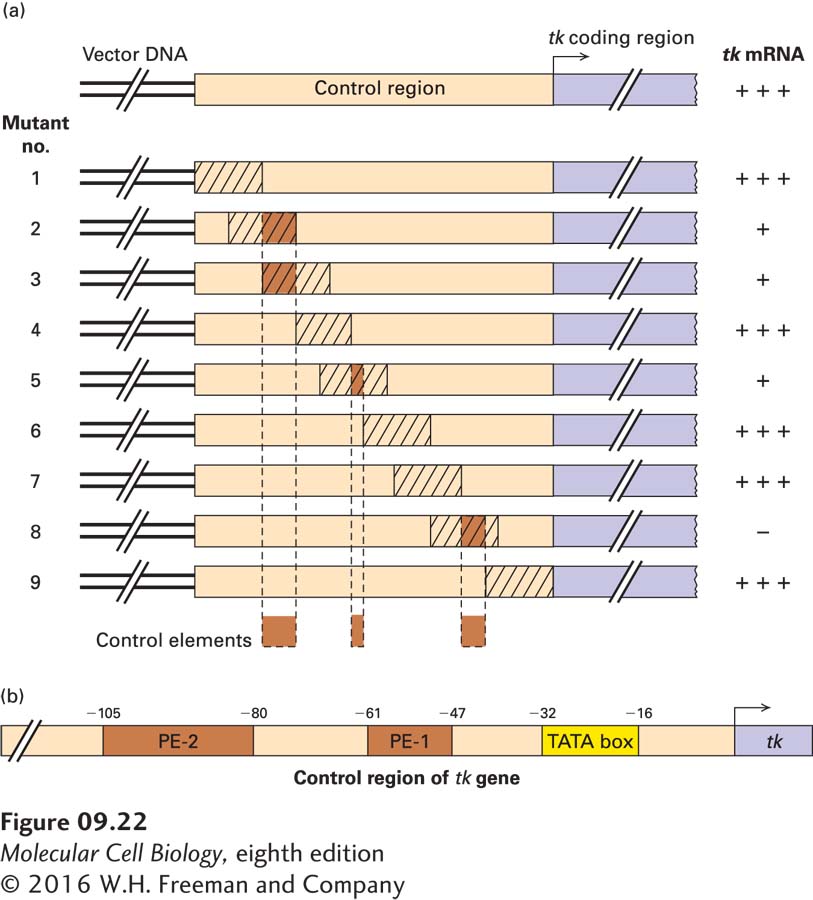
Promoter-Proximal Elements Help Regulate Eukaryotic Genes
Recombinant DNA techniques have been used to systematically mutate the nucleotide sequences of various eukaryotic genes in order to identify transcription-control regions. The use of linker scanning mutagenesis, for example, can pinpoint the sequences within a regulatory region that function to control transcription. In this approach, a set of constructs with contiguous overlapping mutations are assayed for their effect on expression of a reporter gene or production of a specific mRNA (Figure 9-22a). This type of analysis identified promoter-proximal elements of the thymidine kinase (tk) gene from herpes simplex type I virus (HSV-I). The results demonstrated that the DNA region upstream of the HSV-I tk gene contains three separate transcription-control sequences: a TATA box in the interval from −32 to −16 and two other control elements farther upstream (Figure 9-22b). Experiments using mutants containing single-base-pair changes in promoter-proximal control elements revealed that these elements are generally about 6–10 bp long. Recent results indicate that in human genes, they are found both upstream and downstream of the transcription start site at equal frequency. While, strictly speaking, the term promoter refers to the DNA sequence that determines where a polymerase initiates transcription, the term is often used to refer to both a promoter and its associated promoter-proximal control elements.
EXPERIMENTAL FIGURE 9-22Linker scanning mutations identify transcription-control elements. (a) In linker scanning mutagenesis, a region of eukaryotic DNA (tan) that supports high-level expression of a reporter gene (light purple) is cloned in a plasmid vector as diagrammed at the top. Overlapping linker scanning (LS) mutations (crosshatched areas) are introduced from one end of the region being analyzed to the other. These mutations are created by scrambling the nucleotide sequence in a short stretch of the DNA. After the mutant plasmids are transfected separately into cultured cells, the activity of the reporter-gene product is assayed. In the example shown here, the sequence from −120 to +1 of the herpes simplex virus thymidine kinase gene, LS mutations 1, 4, 6, 7, and 9 have little or no effect on expression of the reporter gene, indicating that the regions altered in these mutants contain no control elements. Reporter-gene expression is significantly reduced in mutants 2, 3, 5, and 8, indicating that control elements (brown) lie in the intervals shown at the bottom. (b) Analysis of these LS mutations identified a TATA box and two promoter-proximal elements (PE-1 and PE-2). See S. L. McKnight and R. Kingsbury, 1982, Science217:316.
To test the spacing constraints on control elements in the HSV-I tk promoter region identified by analysis of linker scanning mutations, researchers prepared and assayed constructs containing small deletions and insertions between the elements. Changes in spacing between the promoter and promoter-proximal control elements of 20 bp or fewer had little effect. However, insertions of 30–50 bp between a HSV-I tk promoter-proximal element and the TATA box was equivalent to deleting the element. Similar analyses of other eukaryotic promoters have also indicated that considerable flexibility in the spacing between promoter-proximal elements is generally tolerated, but that separations of several tens of base pairs may decrease transcription.