Activators Are Composed of Distinct Functional Domains
Studies with a yeast transcription activator called Gal4 provided early insight into the domain structure of transcription factors. The gene encoding Gal4, which promotes expression of enzymes needed to metabolize galactose, was identified by complementation analysis of gal4 mutants that cannot form colonies on an agar medium in which galactose is the only source of carbon and energy (see Chapter 6). Directed mutagenesis studies like those described previously identified UASs for the genes activated by Gal4. Each of these UASs was found to contain one or more copies of a 17-bp sequence called UASGAL. DNase I footprinting assays with recombinant Gal4 protein produced in E. coli from the yeast GAL4 gene showed that Gal4 binds to UASGAL sequences. When a copy of UASGAL was cloned upstream of a TATA box followed by a β-galactosidase reporter gene, and that construct was introduced into yeast cells, expression of β-galactosidase was activated in galactose media in wild-type cells, but not in gal4 mutants. These results showed that UASGAL is a transcription-control element activated by the Gal4 transcription factor in galactose media.
Page 382
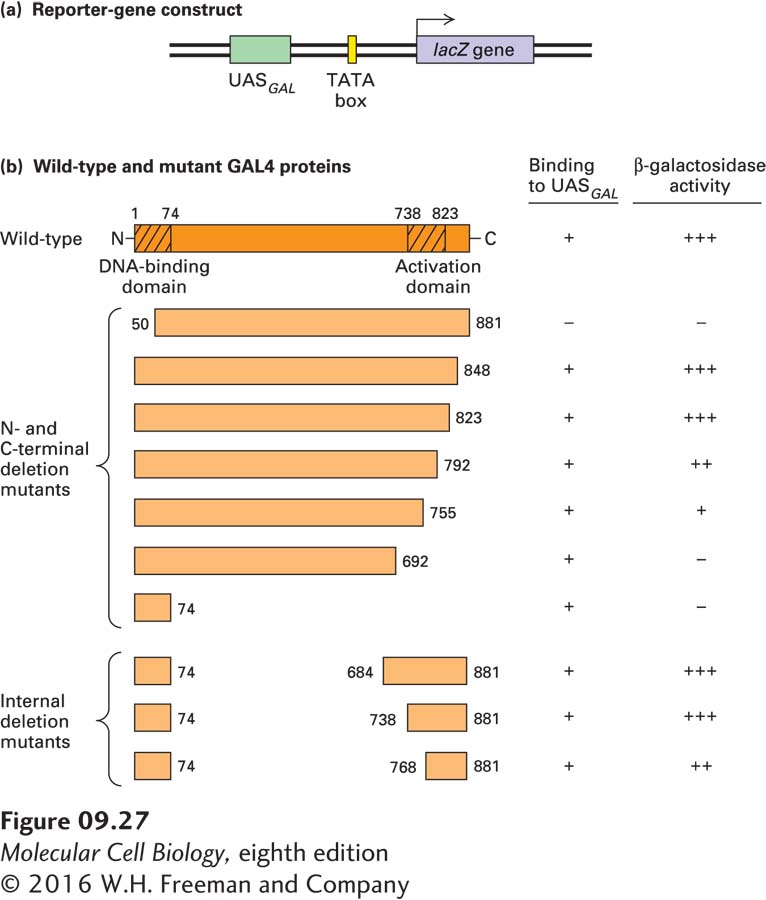
A remarkable set of experiments with gal4 deletion mutants demonstrated that the Gal4 transcription factor is composed of separable functional domains: an N-terminal DNA-binding domain, which binds to specific DNA sequences, and a C-terminal activation domain, which interacts with other proteins to stimulate transcription from a nearby promoter (Figure 9-27). When the N-terminal DNA-binding domain of Gal4 was fused directly to various portions of its own C-terminal region, deleting internal sequences, the resulting truncated proteins retained the ability to stimulate expression of a reporter gene in an in vivo assay like that depicted in Figure 9-26. Thus the internal portion of the protein is not required for the functioning of Gal4 as a transcription factor. Similar experiments with another yeast activator, Gcn4, which regulates genes required for the synthesis of many amino acids, indicated that it contains a roughly 50-amino-acid DNA-binding domain at its C-terminus and a roughly 20-amino-acid activation domain near the middle of its sequence.
EXPERIMENTAL FIGURE 9-27Deletion mutants of the GAL4 gene in yeast with a UASGAL reporter-gene construct demonstrate the separate functional domains in a transcription activator. (a) Diagram of DNA construct containing a lacZ reporter gene (encoding β-galactosidase) and TATA box ligated to UASGAL, a regulatory element that contains several Gal4-binding sites. The reporter-gene construct and DNA encoding wild-type or mutant (deleted) Gal4 were simultaneously introduced into mutant (gal4) yeast cells, and the activity of β-galactosidase expressed from lacZ was assayed. Activity should be high if the introduced GAL4 DNA encodes a functional protein. (b) Schematic diagrams of wild-type Gal4 and various mutant forms. Small numbers refer to positions in the wild-type sequence. Deletion of 50 amino acids from the N-terminal end destroyed the ability of Gal4 to bind to UASGAL and to stimulate expression of β-galactosidase from the reporter gene. Proteins with extensive deletions from the C-terminal end still bound to UASGAL. These results localize the DNA-binding domain to the N-terminal end of Gal4. The ability to activate β-galactosidase expression was not entirely eliminated unless somewhere between 126 and 189 or more amino acids were deleted from the C-terminal end. Thus the activation domain lies in the C-terminal region of Gal4. Proteins with internal deletions (bottom) were also able to stimulate expression of β-galactosidase, indicating that the central region of Gal4 is not crucial for its function in this assay. See J. Ma and M. Ptashne, 1987, Cell48:847; I. A. Hope and K. Struhl, 1986, Cell46:885; and R. Brent and M. Ptashne, 1985, Cell43:729.
Further evidence for the existence of distinct activation domains in Gal4 and Gcn4 came from experiments in which their activation domains were fused to a DNA-binding domain from an entirely unrelated E. coli DNA-binding protein. When these fusion proteins were assayed in vivo, they activated transcription of a reporter gene containing the cognate site for the E. coli protein. Thus functional transcription factors can be constructed from entirely novel combinations of prokaryotic and eukaryotic elements.
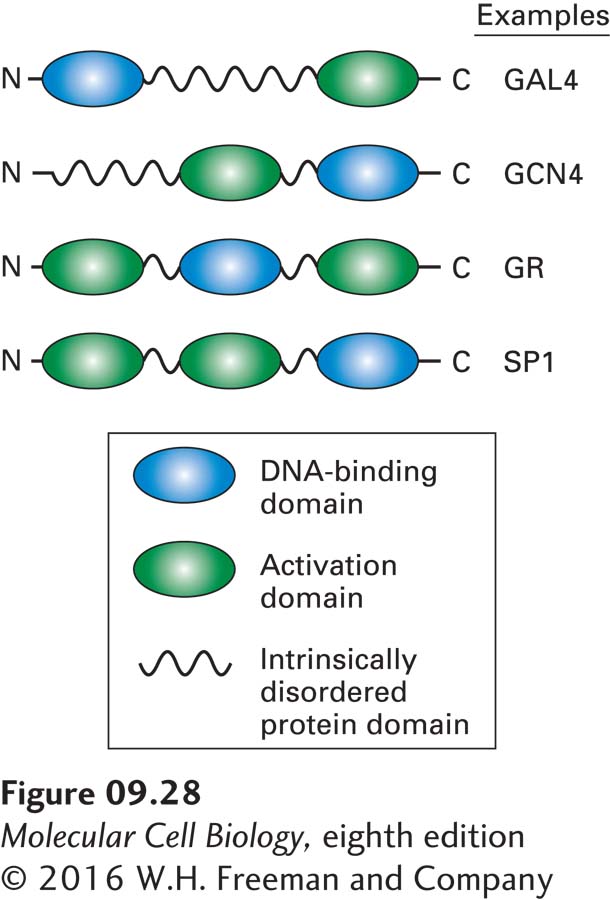
Studies such as these have now been carried out with many eukaryotic transcription factors. The structural model of eukaryotic activators that has emerged from these studies is a modular one in which one or more activation domains are connected to a sequence-specific DNA-binding domain by intrinsically disordered, flexible protein domains (Figure 9-28). In some cases, amino acids included in the DNA-binding domain also contribute to transcriptional activation. As discussed in a later section, activation domains are thought to function by binding other proteins involved in transcription. The presence of flexible, intrinsically disordered protein domains (see Figure 3-8) connecting the DNA-binding domain to the activation domains may explain why alterations in the spacing between control elements are so well tolerated in eukaryotic control regions. Thus even when the positions of transcription factors bound to DNA are shifted relative to each other, their activation domains may still be able to interact because they are attached to their DNA-binding domains through flexible protein regions.
Page 383
FIGURE 9-28Schematic diagrams illustrating the modular structure of eukaryotic transcription activators. Transcription factors may contain more than one activation domain but rarely contain more than one DNA-binding domain. Gal4 and Gcn4 are yeast transcription activators. The glucocorticoid receptor (GR) promotes transcription of target genes when certain hormones are bound to the C-terminal activation domain. SP1 binds to GC-rich promoter elements in a large number of mammalian genes.