Structurally Diverse Activation and Repression Domains Regulate Transcription
Experiments with fusion proteins composed of the Gal4 DNA-binding domain and random segments of E. coli proteins demonstrated that a diverse group of amino acid sequences (~1 percent of all E. coli sequences) can function as activation domains, even though they evolved to perform other functions. Many transcription factors contain activation domains marked by an unusually high percentage of particular amino acids. Gal4, Gcn4, and most other yeast transcription factors, for instance, have activation domains that are rich in acidic amino acids (aspartic and glutamic acids). These so-called acidic activation domains are generally capable of stimulating transcription in nearly all types of eukaryotic cells—fungal, animal, and plant cells. Activation domains from some Drosophila and mammalian transcription factors are glutamine-rich, and some are proline-rich; still others are rich in the closely related amino acids serine and threonine, both of which have hydroxyl groups. However, some strong activation domains are not particularly rich in any specific amino acid.
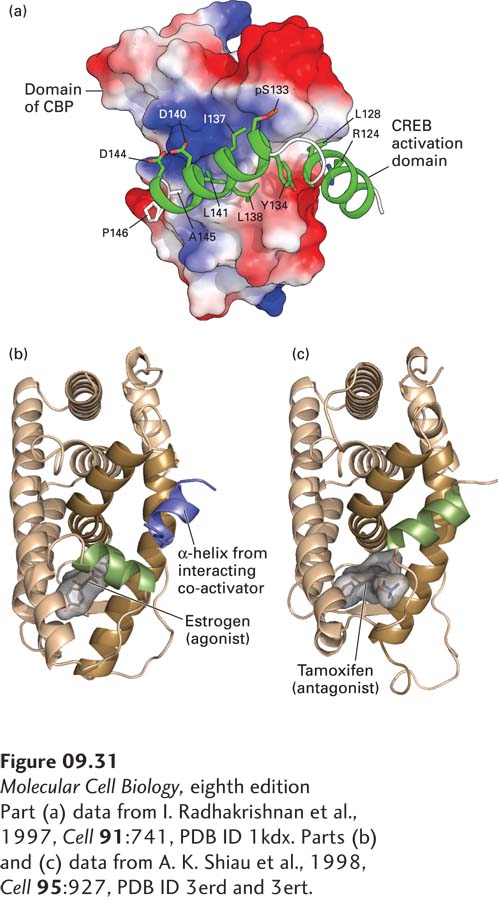
Biophysical studies indicate that acidic activation domains have an unstructured, random-coil, intrinsically disordered conformation. These domains stimulate transcription when they are bound to a protein co-activator. The interaction with a co-activator causes the activation domain to assume a more structured α-helical conformation in the activation domain–co-activator complex. A well-studied example of a transcription factor with an acidic activation domain is the mammalian CREB protein, which is phosphorylated in response to increased levels of cAMP. This regulated phosphorylation is required for CREB to bind to its co-activator CBP (CREB binding protein), resulting in the transcription of genes whose control regions contain a CREB-binding site (see Figure 15-30). When the phosphorylated random-coil activation domain of CREB interacts with CBP, it undergoes a conformational change to form two α helices linked by a short loop, which wrap around the interacting domain of CBP (Figure 9-31a).
[Part (a) data from I. Radhakrishnan et al., 1997, Cell91:741, PDB ID 1kdx. Parts (b) and (c) data from A. K. Shiau et al., 1998, Cell95:927, PDB ID 3erd and 3ert.]
FIGURE 9-31Activation domains may be random coils until they interact with co-activator proteins or folded protein domains. (a) The acidic activation domain of CREB (cyclic AMP response element-binding protein) is activated by phosphorylation at serine 123. It is a random coil until it interacts with a domain of its co-activator, CBP (shown as a space-filling surface model with negatively charged regions in red and positively charged regions in blue). When the CREB activation domain binds to CBP, it folds into two amphipathic α helices. Side chains in the activation domain that interact with the surface of the CBP domain are labeled. (b) The ligand-binding activation domain of the estrogen receptor is a folded-protein domain. When estrogen is bound to the domain, the green α helix interacts with the ligand, generating a hydrophobic groove in the ligand-binding domain (dark brown helices), which binds an amphipathic α helix in a co-activator subunit (blue). (c) The conformation of the estrogen receptor in the absence of hormone is stabilized by binding of the estrogen antagonist tamoxifen. In this conformation, the green helix of the receptor folds into a conformation that interacts with the co-activator–binding groove of the active receptor, sterically blocking binding of co-activators.
[Part (a) data from I. Radhakrishnan et al., 1997, Cell91:741, PDB ID 1kdx. Parts (b) and (c) data from A. K. Shiau et al., 1998, Cell95:927, PDB ID 3erd and 3ert.]
Page 387
Some activation domains are larger and more highly structured than acidic activation domains. For example, the ligand-binding domains of nuclear receptors function as activation domains when they bind their specific hormone ligand (Figure 9-31b, c). Binding of ligand induces a large conformational change in the nuclear receptor that allows the ligand-binding domain with bound hormone to interact with a short α helix in a co-activator; the resulting complex can then activate transcription of genes whose control regions bind the nuclear receptor.
Thus the acidic activation domain in CREB and the ligand-binding activation domains in nuclear receptors represent two structural extremes. The CREB acidic activation domain is an intrinsically disordered random coil that folds into two α helices when it binds to the surface of a globular domain in a co-activator. In contrast, the nuclear-receptor ligand-binding activation domain is a structured globular domain that interacts with a short α helix in a co-activator, which probably is a random coil before it is bound. In both cases, however, specific protein-protein interactions between a co-activator and the activation domain permit the transcription factor to stimulate gene expression.
Currently, less is known about the structure of repression domains. The globular ligand-binding domains of some nuclear receptors function as repression domains in the absence of their specific hormone ligand. Like activation domains, repression domains may be relatively short, comprising 15 or fewer amino acids. Biochemical and genetic studies indicate that repression domains also mediate protein-protein interactions and bind to co-repressor proteins, forming a complex that inhibits transcription initiation by mechanisms that are discussed later in the chapter.