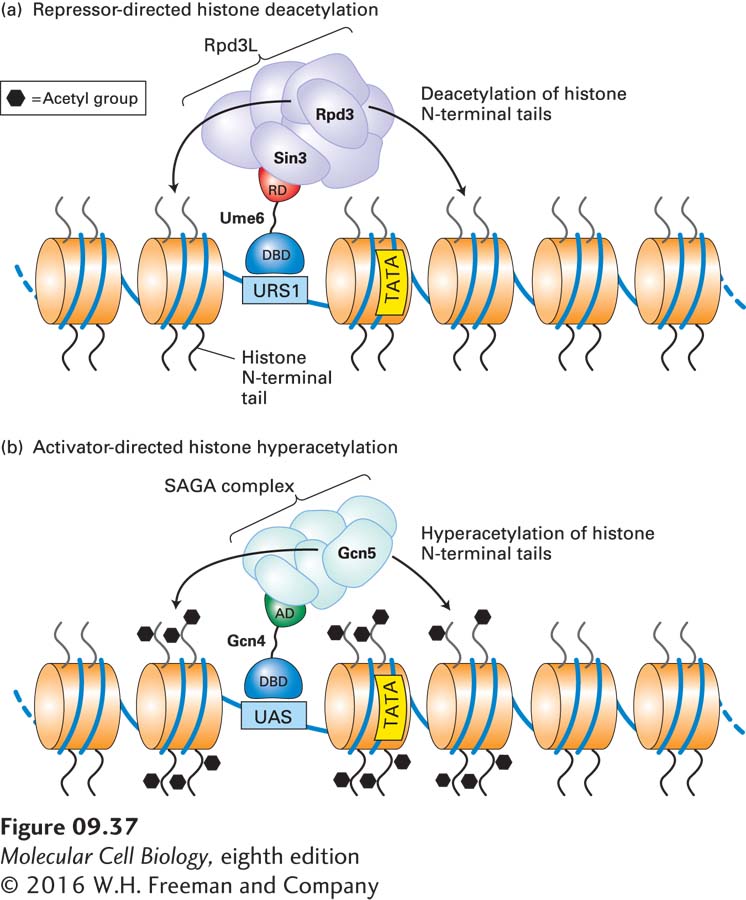
Repressors Can Direct Histone Deacetylation at Specific Genes
The importance of histone deacetylation in chromatin-mediated gene repression was further supported by studies of eukaryotic repressors that regulate genes at internal chromosomal positions. These proteins are now known to act in part by causing deacetylation of histone tails in nucleosomes that encompass the TATA box and promoter-proximal region of the genes they repress. In vitro studies have shown that when promoter DNA is part of a nucleosome with nonacetylated histones, the general transcription factors cannot bind to the TATA box and promoter-proximal region. In nonacetylated histones, the N-terminal lysines are positively charged and may interact with DNA phosphates. The nonacetylated histone tails also interact with neighboring histone octamers and other chromatin-associated proteins, favoring the folding of chromatin into condensed higher-order structures whose precise conformation is not well understood. The net effect is that general transcription factors cannot assemble into a preinitiation complex on a promoter associated with hypoacetylated histones. In contrast, binding of general transcription factors is repressed much less by histones with hyperacetylated tails, in which the positively charged lysines are neutralized and electrostatic interactions are eliminated.
The connection between histone deacetylation and repression of transcription at specific yeast promoters became clearer when the cDNA encoding a human histone deacetylase was found to have high homology to the yeast RPD3 gene, known to be required for the normal repression of a number of yeast genes. Further work showed that the yeast Rpd3 protein has histone deacetylase activity. The ability of Rpd3 to deacetylate histones at a number of promoters depends on two other proteins: Ume6, a repressor that binds to a specific upstream regulatory sequence (URS1), and Sin3, which is part of a large multiprotein complex called Rpd3L that also contains Rpd3 (Figure 9-37a). Sin3 also binds to the repression domain of Ume6, thus positioning the Rpd3 histone deacetylase in the complex so that it can interact with nearby promoter-associated nucleosomes and remove acetyl groups from histone-tail lysines. Additional experiments, using the chromatin immunoprecipitation technique outlined in Figure 9-18a and antibodies to specific histone acetylated lysines, demonstrated that in wild-type yeast, one or two nucleosomes in the immediate vicinity of Ume6-binding sites are hypoacetylated. These sites include the promoters of genes repressed by Ume6. In sin3 and rpd3 deletion mutants, not only were these promoters derepressed, but the nucleosomes near the Ume6-binding sites were hyperacetylated. All these findings provide considerable support for the model of repressor-directed deacetylation shown in Figure 9-37a.
FIGURE 9-37Proposed mechanism of histone deacetylation and hyperacetylation in yeast transcriptional control. (a) Repressor-directed deacetylation of histone N-terminal tails. The DNA-binding domain (DBD) of the repressor Ume6 interacts with a specific upstream control element of the genes it regulates, called URS1. The Ume6 repression domain (RD) binds Sin3, a subunit of a multiprotein complex that includes Rpd3, a histone deacetylase. Deacetylation of histone N-terminal tails on nucleosomes in the region of the Ume6-binding site inhibits binding of general transcription factors at the TATA box, thereby repressing gene expression. (b) Activator-directed hyperacetylation of histone N-terminal tails. The DNA-binding domain of the activator Gcn4 interacts with specific upstream activating sequences (UAS) of the genes it regulates. The Gcn4 activation domain (AD) then interacts with a multiprotein histone acetylase complex that includes the Gcn5 catalytic subunit. Subsequent hyperacetylation of histone N-terminal tails on nucleosomes in the vicinity of the Gcn4-binding site facilitates access by the general transcription factors required for initiation. Repression and activation of many genes in higher eukaryotes occur by similar mechanisms.
In yeast, the Sin3-Rpd3 complex (Rpd3L) functions as a co-repressor, a protein or complex of proteins that binds to a repression domain and interacts with chromatin, Pol II, or the general transcription factors to repress transcription. Co-repressor complexes containing histone deacetylases have also been found associated with many repressors from mammalian cells. Some of these complexes contain the mammalian homolog of Sin3 (mSin3), which interacts with the repression domain of repressors, as in yeast. Other histone deacetylase complexes identified in mammalian cells contain additional or different repression domain-binding proteins. These various repressor and co-repressor combinations mediate histone deacetylation at specific promoters by a mechanism similar to the yeast mechanism (see Figure 9-37a). In addition to repressing transcription through the formation of “closed” chromatin structures, some repression domains have also been found to inhibit the assembly of preinitiation complexes in in vitro experiments with purified general transcription factors in the absence of histones. This activity probably contributes to the repression of transcription by these repression domains in vivo as well.