The Mediator Complex Forms a Molecular Bridge Between Activation Domains and Pol II
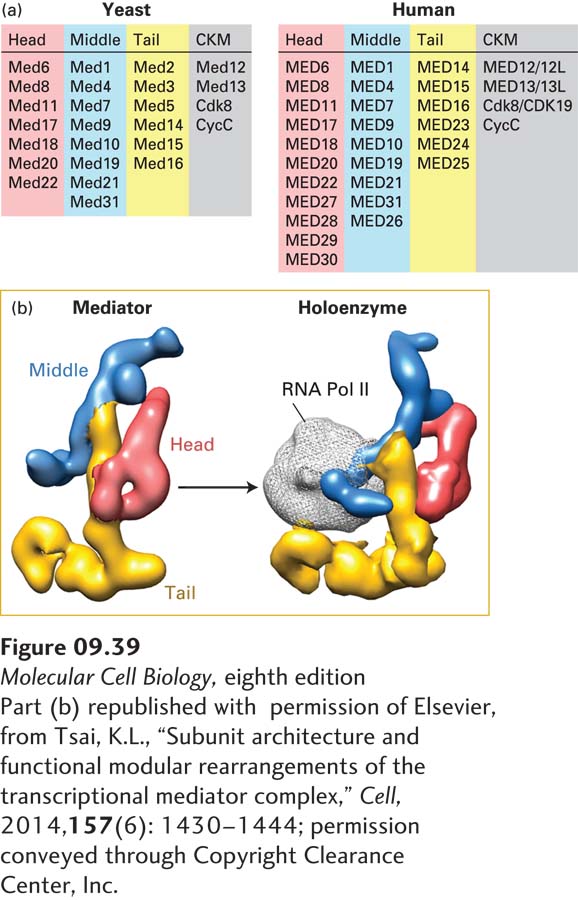
Once the interaction of activation domains with histone acetylase complexes and chromatin remodeling complexes converts the chromatin of a promoter region to an “open” structure that allows the binding of general transcription factors, activation domains interact with another multisubunit co-activator complex, the Mediator complex (Figure 9-39). Activation domain–Mediator interactions stimulate assembly of the preinitiation complex on the promoter. Recent cryoelectron microscopy studies show that the head and middle domains of the Mediator complex interact directly with Pol II. Several Mediator subunits bind to activation domains in various activator proteins. Thus Mediator can form a molecular bridge between an activator bound to its cognate site in DNA and Pol II bound to a promoter.
[Part (b) republished with permission of Elsevier, from Tsai, K.L., “Subunit architecture and functional modular rearrangements of the transcriptional mediator complex,” Cell, 2014, 157(6): 1430–1444; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 9-39Structure of yeast and human Mediator complexes. (a) Subunits of the S. cerevisiae and human Mediator complexes. The subunits constituting the head, middle, and tail modules of Mediator are indicated, as well as the subunits of the CDK8-kinase module (CKM) that associates with some Mediator complexes, blocking Pol II binding. (b) Cryoelectron microscopic structure of the yeast Mediator without the CKM. (Left) The head, middle, and tail modules composed of the subunits listed above are color-coded. (Right) The structure of a complex of Mediator with Pol II, called the holoenzyme, suggests that the Mediator modules rotate relative to one another as shown to create a surface that binds Pol II.
[Part (b) republished with permission of Elsevier, from Tsai, K.L., “Subunit architecture and functional modular rearrangements of the transcriptional mediator complex,” Cell, 2014, 157(6): 1430–1444; permission conveyed through Copyright Clearance Center, Inc.]
Experiments with temperature-sensitive yeast mutants indicate that some Mediator subunits are required for transcription of virtually all yeast genes. These subunits help maintain the overall structure of the Mediator complex or bind to Pol II; they are therefore required for activation by all activators. In contrast, other Mediator subunits are required for normal activation or repression of specific subsets of genes. DNA microarray analysis (see Figure 6-26) of yeast gene expression in mutants with defects in these nonessential Mediator subunits have indicated that each one influences transcription of 3–10 percent of all genes to the extent that its deletion either increases or decreases mRNA expression by a factor of twofold or more. In many cases, these Mediator subunits have been discovered to interact with specific activation domains; thus when one Mediator subunit is defective, transcription of genes regulated by activators that bind to that subunit is severely depressed, but transcription of other genes is unaffected. Recent cryoelectron microscopy studies suggest that when activation domains interact with Mediator, the head, middle, and tail domains depicted in Figure 9-39 rotate relative to one another, creating a binding surface for RNA polymerase II. The surface of the polymerase that interacts with general transcription factors in the preinitiation complex (see Figure 9-20) remains exposed in the proposed model of the polymerase-Mediator complex, referred to as the holoenzyme.
Page 397
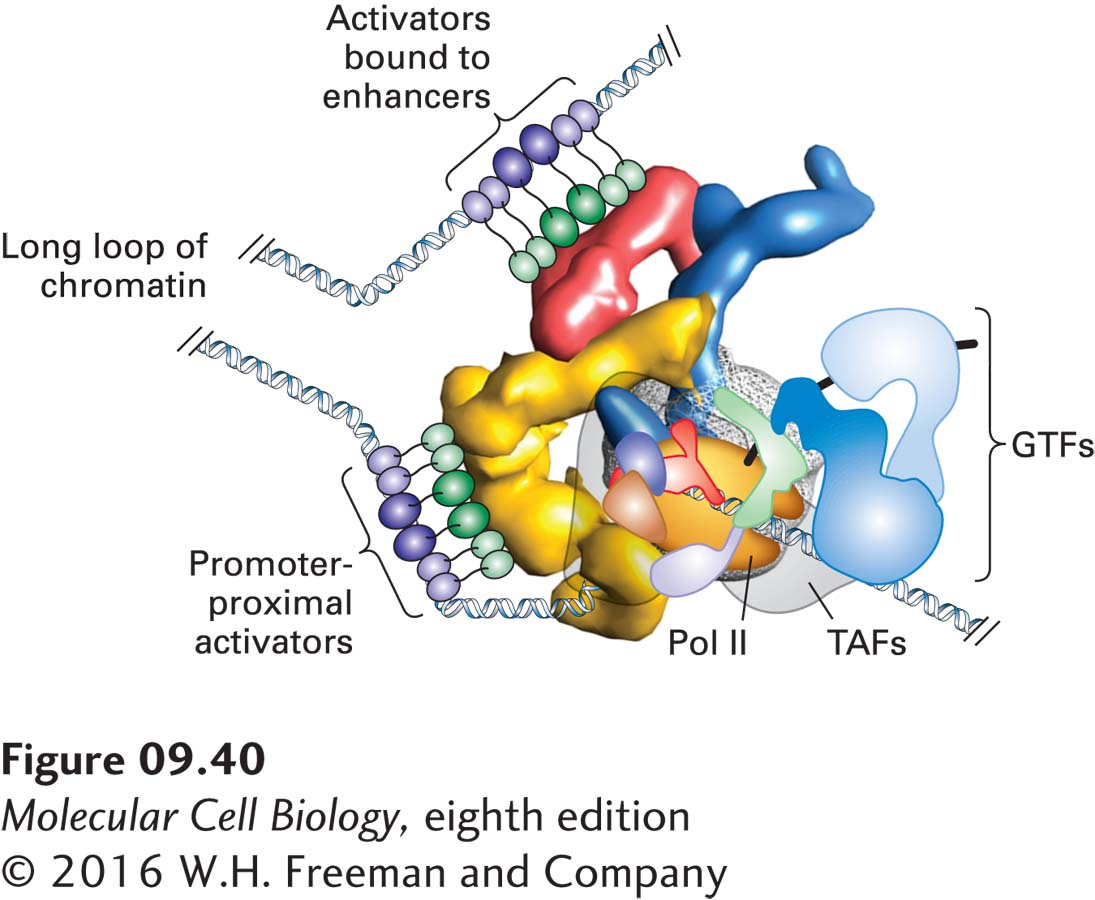
The various experimental results indicating that individual Mediator subunits bind to specific activation domains suggest that multiple activators may influence transcription from a single promoter by interacting with a Mediator complex simultaneously or in rapid succession (Figure 9-40). Activators bound at enhancers or promoter-proximal elements can interact with Mediator associated with a promoter because chromatin, like DNA, is flexible and can form a loop, bringing the regulatory regions and the promoter close together, as observed for the E. coli NtrC activator and σ54-RNA polymerase (see Figure 9-5). The multiprotein complexes that form on eukaryotic promoters may comprise more than 100 polypeptides with a total mass of 3–5 megadaltons (MDa)—as large as a ribosome.
FIGURE 9-40Model of several DNA-bound activators interacting with a single Mediator complex. The ability of different Mediator subunits to interact with specific activation domains may contribute to the integration of signals from several activators at a single promoter. See the text for discussion.
In vivo, assembly of a preinitiation complex on a promoter and initiation of transcription is a highly cooperative process generally requiring that several transcription factors bound to transcription-control elements interact with co-activators that in turn interact with Pol II and general transcription factors. A cell must produce the specific set of activators required for transcription of a particular gene in order to express that gene.