Expression of Many Bacterial Operons Is Controlled by Regulation of Transcriptional Elongation
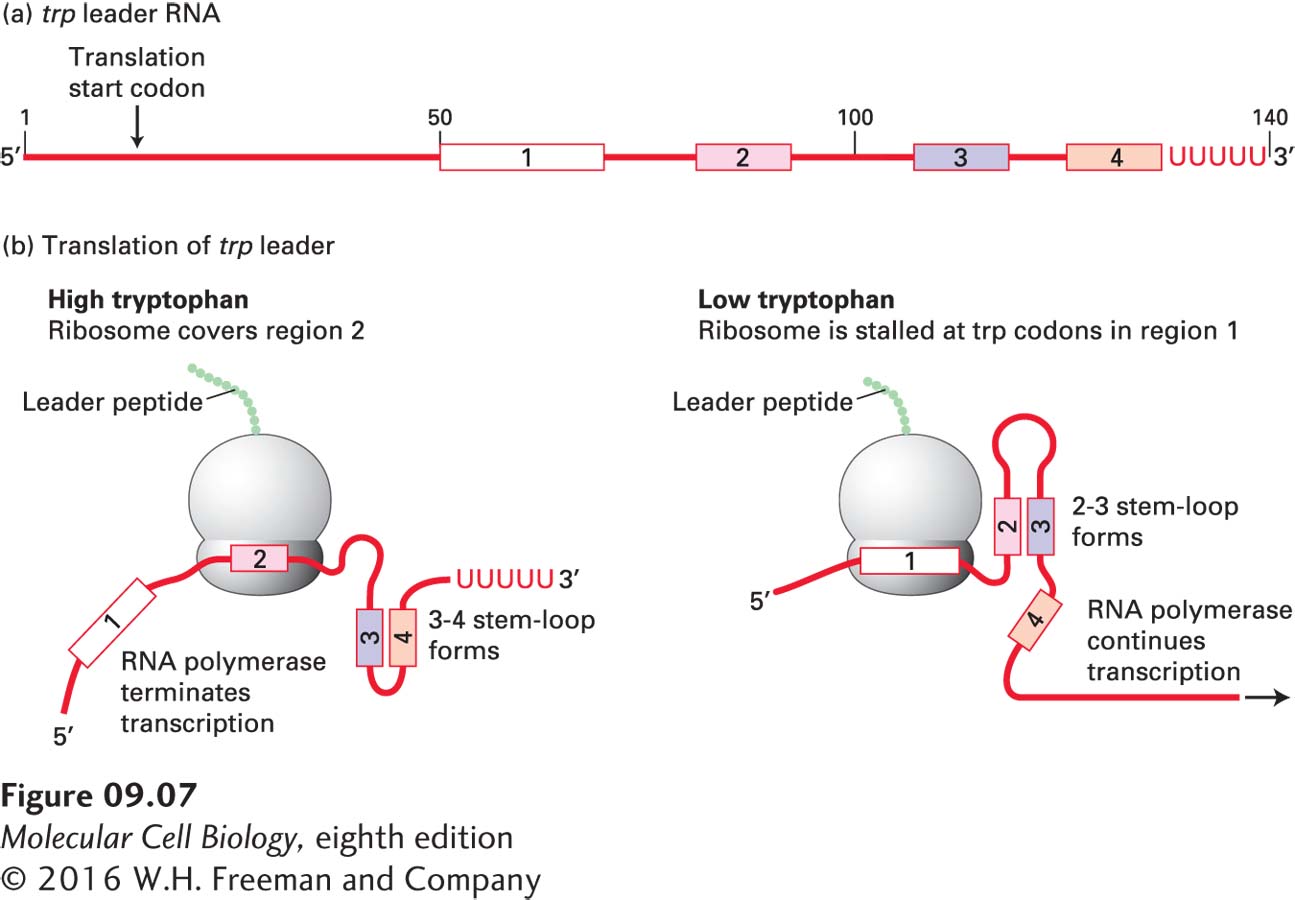
In addition to regulation of transcription initiation by activators and repressors, expression of many bacterial operons is controlled by regulation of transcriptional elongation in the promoter-proximal region. This mechanism of control was first discovered in studies of trp operon transcription in E. coli (see Figure 5-13). Transcription of the trp operon is repressed by the trp repressor when the concentration of tryptophan in the cytoplasm is high. But the low level of transcription initiation that still occurs is further controlled by a process called attenuation when the concentration of charged tRNATrp is sufficient to support a high rate of protein synthesis. The first 140 nt of the trp operon does not encode proteins required for tryptophan biosynthesis, but rather consists of a short peptide “leader sequence,” as diagrammed in Figure 9-7a. Region 1 of this leader sequence contains two successive Trp codons. Region 3 can base-pair with either region 2 or region 4. A ribosome follows closely behind the RNA polymerase, initiating translation of the leader peptide shortly after the 5′ end of the trp leader sequence emerges from the RNA polymerase. When the concentration of tRNATrp is sufficient to support a high rate of protein synthesis, the ribosome translates quickly through region 1 into region 2, blocking the ability of region 2 to base-pair with region 3 as it emerges from the surface of the transcribing RNA polymerase (Figure 9-7b, left). Instead, region 3 base-pairs with region 4 as soon as it emerges from the surface of the polymerase, forming a stem-loop (see Figure 5-9a) followed by several uracils, which is a signal for bacterial RNA polymerase to pause transcription and terminate. As a consequence, the remainder of the long trp operon is not transcribed, and the cell does not waste the energy required for tryptophan synthesis, or for the translation of the encoded proteins, when the concentration of tryptophan is high.
FIGURE 9-7Transcriptional control by regulation of RNA polymerase elongation and termination in the E. coli trp operon. (a) Diagram of the 140-nucleotide trp leader RNA. The numbered regions are critical to attenuation. (b) Translation of the trp leader sequence begins near the 5′ end soon after it is transcribed, while transcription of the rest of the polycistronic trp mRNA molecule continues. At high concentrations of charged tRNATrp, formation of the 3–4 stem-loop followed by a series of uracils causes termination of transcription. At low concentrations of charged tRNATrp, region 3 is sequestered in the 2–3 stem-loop and cannot base-pair with region 4. In the absence of the stem-loop structure required for termination, transcription of the trp operon continues. See C. Yanofsky, 1981, Nature289:751.
Page 362
However, when the concentration of tRNATrp is not sufficient to support a high rate of protein synthesis, the ribosome stalls at the two successive Trp codons in region 1 (Figure 9-7b, right). As a consequence, region 2 base-pairs with region 3 as soon as it emerges from the transcribing RNA polymerase. This prevents region 3 from base-pairing with region 4, so the 3–4 hairpin does not form and does not cause RNA polymerase pausing or transcription termination. As a result, the proteins required for tryptophan synthesis are translated by ribosomes that initiate translation at the start codons for each of these proteins in the long polycistronic trp mRNA.
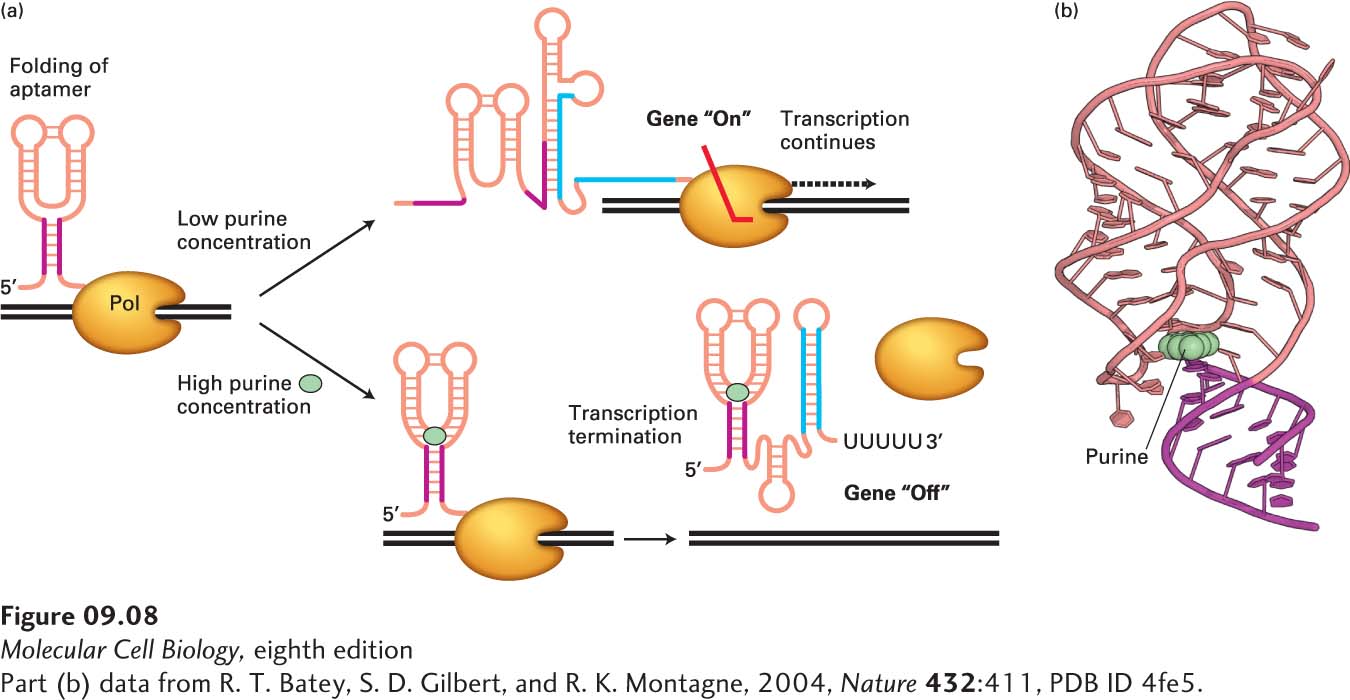
Attenuation of transcription elongation also occurs at some operons and single genes encoding enzymes involved in the biosynthesis of other amino acids and metabolites through the function of riboswitches. Riboswitches are sequences of RNA most commonly found in the 5′ untranslated region of bacterial mRNAs. They fold into complex tertiary structures called aptamers that bind small-molecule metabolites when those metabolites are present at sufficiently high concentrations. In some cases, this binding results in the formation of stem-loop structures that lead to early termination of transcription, as in the Bacillus subtilis xpt-pbuX operon, which encodes enzymes involved in purine synthesis (Figure 9-8). When the concentration of small-molecule metabolites is lower, the metabolites are not bound by the aptamers, and alternative RNA structures form that do not induce transcription termination, allowing transcription of genes encoding enzymes involved in the synthesis of the metabolites. As we will see below, although the mechanism in eukaryotes is different, regulation of promoter-proximal transcriptional pausing and termination has recently been discovered to occur frequently in the regulation of gene expression in multicellular organisms as well.
[Part (b) data from R. T. Batey, S. D. Gilbert, and R. K. Montagne, 2004, Nature432:411, PDB ID 4fe5.]
FIGURE 9-8Riboswitch control of transcription termination in B. subtilis. (a) During transcription of the Bacillus subtilis xpt-pbuX operon, which encodes enzymes involved in purine synthesis, the 5′ untranslated region of the mRNA can fold into alternative structures depending on the concentration of purines in the cytoplasm, forming the “purine riboswitch.” At high concentrations of purines, the riboswitch folds into an aptamer that binds a purine ligand (cyan circle), allowing formation of a stem-loop transcription termination signal similar to the termination signal that forms in the E. coli trp operon mRNA at high tryptophan concentrations (see Figure 9-7), i.e., a stem loop followed by a run of Us. At low purine concentrations, an alternative RNA structure forms that prevents formation of the transcription termination signal, permitting transcription of the operon. Note the alternative base pairing of the red and blue regions of the RNA. (b) Structure of the purine riboswitch bound to a purine (cyan) as determined by X-ray crystallography. See A. D. Garst, A. L. Edwards, and R. T. Batey, 2011, Cold Spring Harb. Perspect. Biol.3:a003533.
[Part (b) data from R. T. Batey, S. D. Gilbert, and R. K. Montagne, 2004, Nature432:411, PDB ID 4fe5.]