
Chapter 9. Coordination of Eukaryotic Polymerases
Introduction
Analyze the Data 9-1: Coordination of Eukaryotic Polymerases
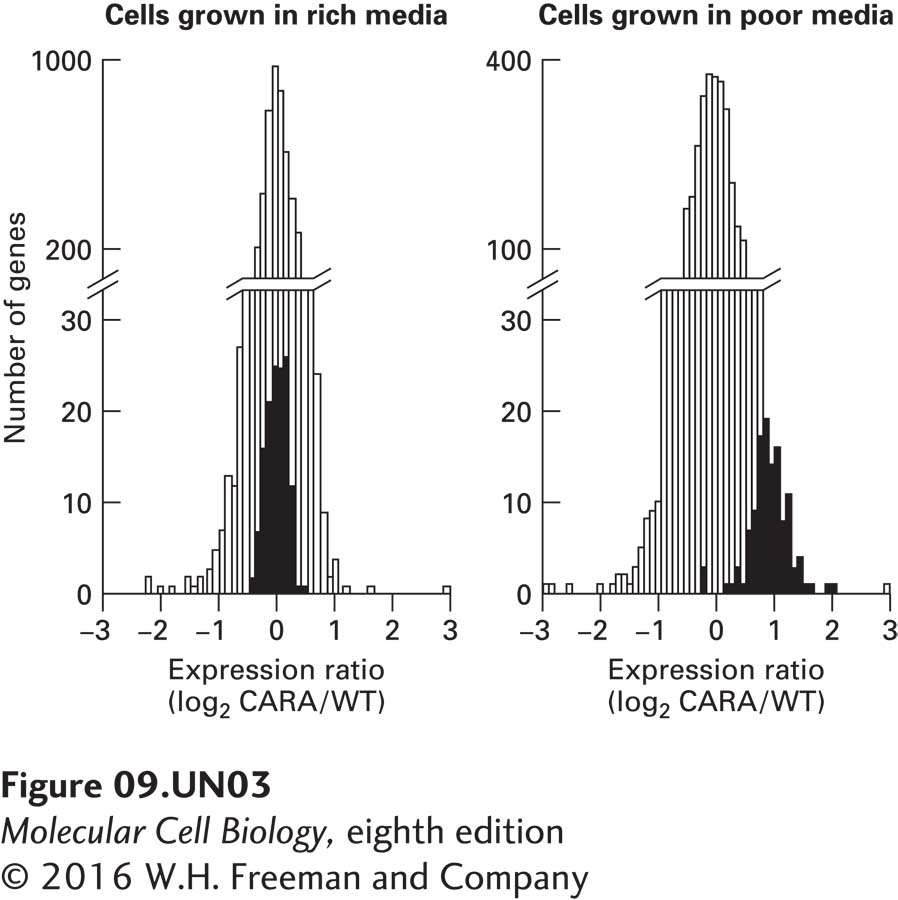
In eukaryotes, the three RNA polymerases, Pol I, II, and III, each transcribe unique genes required for the synthesis of ribosomes: 25S and 18S rRNAs (Pol I), 5S rRNA (Pol III), and mRNAs for ribosomal proteins (Pol II). Researchers have long speculated that the activities of the three RNA polymerases are coordinately regulated according to the demand for ribosome synthesis: high demand in replicating cells in rich nutrient conditions and low demand when nutrients are scarce. To determine whether the activities of the three polymerases are coordinated, Arnaud Laferte and colleagues engineered a strain of yeast to be partially resistant to the inhibition of cell growth by the drug rapamycin (2006, Genes Dev. 20:2030–2040). As discussed in Chapter 10, rapamycin inhibits a protein kinase called TOR (for target of rapamycin) that regulates the overall rate of protein synthesis and ribosome synthesis. When TOR is inhibited by rapamycin, the transcription of rRNAs by Pol I and Pol III and of ribosomal protein mRNAs by RNA polymerase II are all rapidly repressed. Part of the inhibition of rRNA synthesis results from the dissociation of the Pol I transcription factor Rrn3 from Pol I. In the strain constructed by Laferte and colleagues, the wild-type Rrn3 gene and the wild-type A43 gene, encoding the Pol I subunit to which Rrn3 binds, were replaced with a gene encoding a fusion of the A43 Pol I subunit with Rrn3. The idea was that the covalent fusion of the two proteins would prevent the Rrn3 dissociation from Pol I otherwise caused by rapamycin treatment. The resulting CARA (constitutive association of Rrn3 and A43) yeast strain was found to be partially resistant to rapamycin. In the absence of rapamycin, the CARA strain grew at the same rate and had as many ribosomes as wild-type cells.
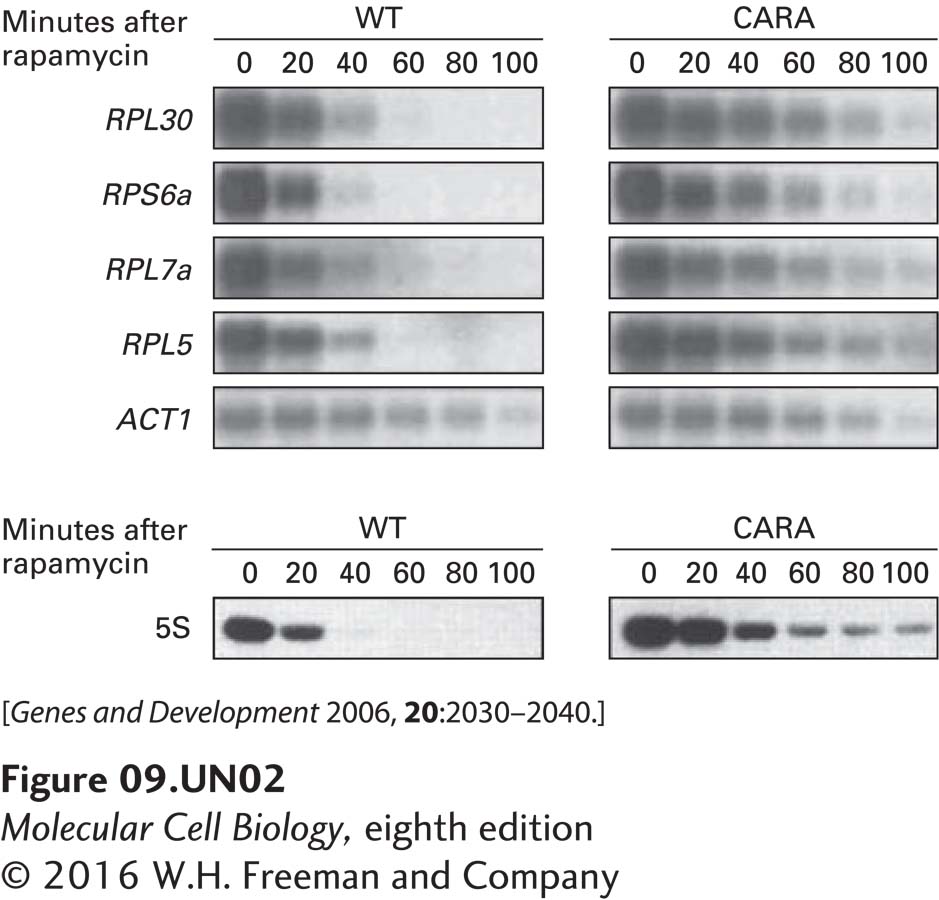
a. To analyze rRNA transcription by Pol I, total RNA was isolated from rapidly growing wild-type (WT) cells and from CARA cells at various times following the addition of rapamycin. The concentration of the 35S rRNA precursor transcribed by Pol I (see Figure 10-41) was assayed by the primer-extension method. Since the 5′ end of the 35S rRNA precursor is degraded during the processing of 28S and 18S rRNA, this method measures the relatively shortlived pre-rRNA precursor. It is therefore an indirect measure of the rate of rRNA transcription by Pol I. The results of this primer-extension assay are shown below. How does the CARA strain’s Pol I–Rrn3 fusion affect the response of Pol I transcription to rapamycin?

In untreated cells (t = 0 minutes), both wild-type and CARA cells transcribed equivalent amounts of ribosomal RNAs, suggesting that the mutant Pol I functions normally under control conditions. This finding is consistent with the observation that the growth rate of yeast harboring this mutation was indistinguishable from wild-type cells. However, when the drug rapamycin was added, Pol I transcription in wild-type cells was completely inhibited by 100 minutes after addition of the drug. In contrast, in the CARA cells, Pol I transcription was much less inhibited and continued at a significant level even at 100 minutes after addition of rapamycin. Consequently, Pol I transcription in the CARA cells was only partially inhibited by rapamycin.
Activity results are being submitted...