Genes in the X chromosome exhibit a “crisscross” inheritance pattern.
When Morgan did his crosses with the white-eyed male, the X chromosome had only recently been discovered by microscopic examination of the chromosomes in male and female grasshoppers. Morgan was the first to understand that the pattern of inheritance of the X chromosome would be different from that of the autosomes, and he proposed the hypothesis that the white-eyed phenotype was due to a mutation in a gene in the X chromosome. This hypothesis could explain the pattern of inheritance shown in Fig. 17.3.
The key features of X-linked inheritance are shown in Fig. 17.4. In Drosophila, as in humans, females are XX and males are XY. Fig. 17.4a shows an XY male in which the X chromosome contains a recessive mutation. Because the Y chromosome does not carry an allele of this gene, the recessive mutation will be reflected in the male’s phenotype—in this case, white eyes.
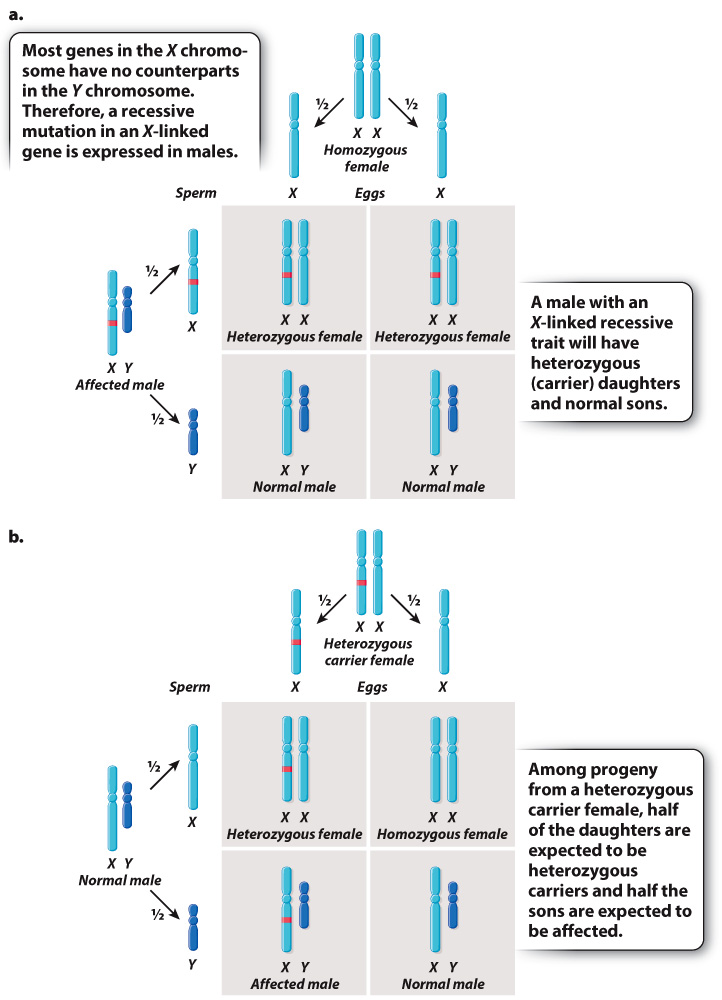
FIG. 17.4 X-linkage. (a) Cross between a homozygous nonmutant female and a male carrying an X-linked recessive allele (red). (b) Cross between a heterozygous female and a nonmutant male. X-linked recessive alleles are expressed in males because males have only one X chromosome.
The Punnett square in Fig. 17.4a explains why all the offspring had red eyes when Morgan crossed the white-eyed male with a red-eyed female in the parental generation. During meiosis in the male, the mutant X chromosome segregates from the Y chromosome, and each type of sperm, X or Y, combines with a normal X-bearing egg. The result is that the female progeny are heterozygous. They have only one copy of the mutant allele, and because the mutant allele is recessive, the heterozygous females do not express the mutant white-eye trait. The male progeny are also red-eyed because they receive their X chromosome from their wild-type red-eyed mother.
The Punnett square in Fig. 17.4a illustrates two important principles governing the inheritance of X-linked genes:
- The phenotypes of the XX offspring indicate that a male transmits his X chromosome only to his daughters. In this case, the X chromosome transmitted by the male carries the white-eye mutation.
Page 349
The phenotypes of the XY offspring indicate that a male inherits his X chromosome from his mother. In this case, the X chromosome transmitted by the mother carries the nonmutant allele of the gene.
These principles underlie a pattern often referred to as crisscross inheritance: An X chromosome present in a male in one generation must be transmitted to a female in the next generation, and in the generation after that can be transmitted back to a male. Therefore, an X chromosome can “crisscross,” or alternate, between the sexes in successive generations.
The mating illustrated in Fig. 17.4b shows another important feature of X-linked inheritance, one that explains the results of Morgan’s F1 cross. In this case, the mother is a heterozygous female. During meiosis, the mutant allele and the nonmutant allele undergo segregation. Half of the resulting eggs contain an X chromosome with the mutant allele, and half contain an X chromosome with the nonmutant allele. These combine at random with either X-bearing sperm or Y-bearing sperm. Among the female progeny in the next generation, half are heterozygous for the mutant allele and the other half are homozygous for the normal allele. Thus, none of the females exhibits the white-eye trait because the mutation is recessive. Among the male progeny, half receive the mutant allele and have white eyes, whereas the other half receive the nonmutant allele and have red eyes. The expected progeny from the cross in Fig. 17.4b therefore consist of all red-eyed females, and there is a 1:1 ratio of red-eyed to white-eyed males, which is what Morgan observed in the F2 generation of his crosses, as shown in Fig. 17.3.
Knowing the patterns revealed by the Punnett squares, we can now assign genotypes to Morgan’s original crosses (Fig. 17.5). In Fig. 17.5, the white-eyed males that Morgan used in his parental generation crosses are given the genotype of w–Y, and the red-eyed female has a genotype of w+w+. The symbol “w–” stands for the recessive white-eye mutation in one X chromosome, and the symbol “w+” stands for the dominant nonmutant allele (red eyes) in the other X chromosome. The symbol “Y” stands for the Y chromosome, and it is important to remember that the Y chromosome does not contain an allele of the white-eye gene.
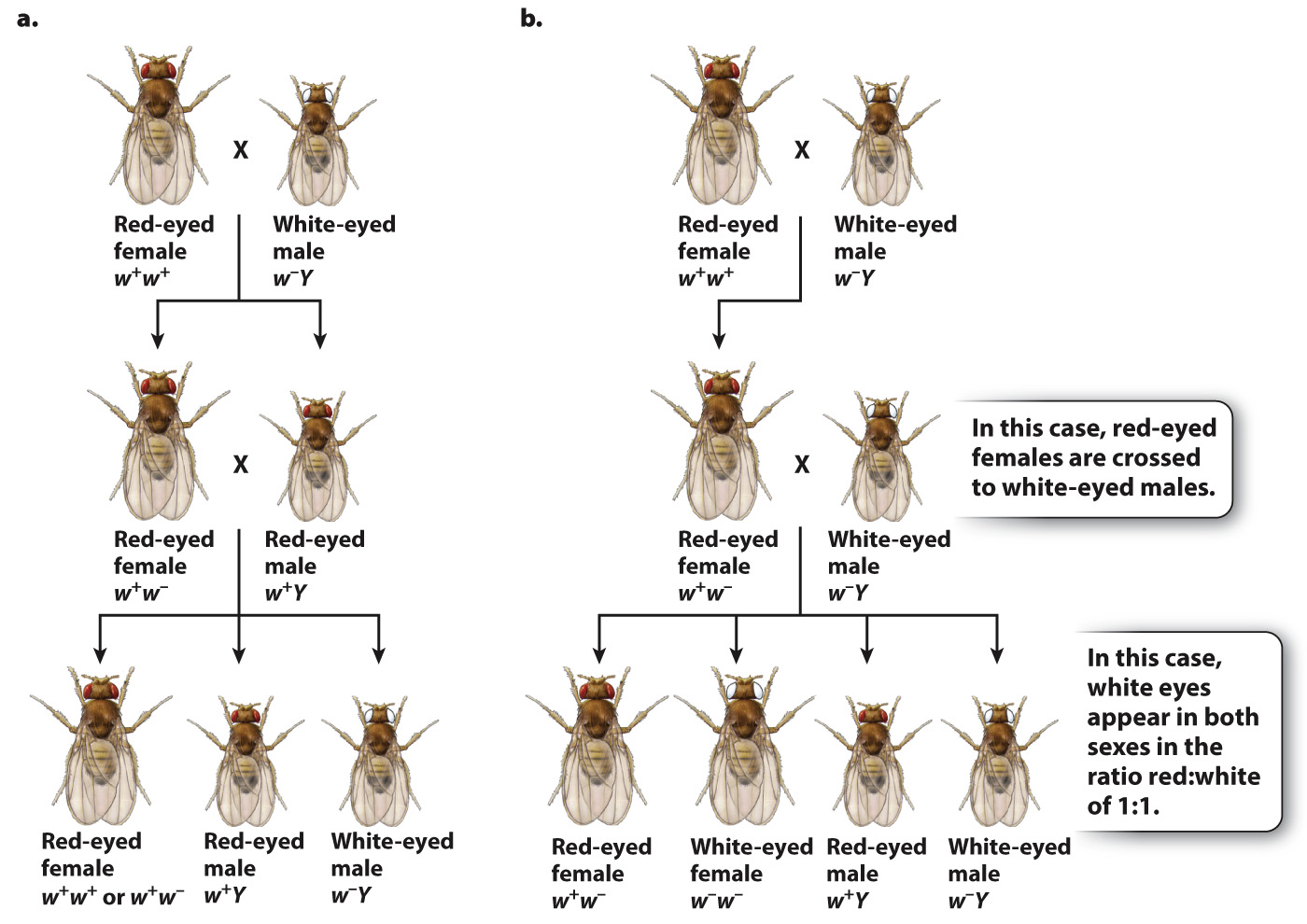
FIG. 17.5 Genotypes of Morgan’s white-eyed fruit fly crosses.
In the cross between a wild-type red-eyed female and a white-eyed male, illustrated in Fig. 17.5a, the male offspring have red eyes because they receive their X chromosome from their mother. The female offspring from this cross receive one of their X chromosomes from their father, and hence they are heterozygous, w+w−. When the male and female progeny are mated together, their offspring consist of all red-eyed females (half of which are heterozygous) and a 1:1 ratio of red-eyed to white-eyed males, exactly as Morgan had observed.
Page 350
The hypothesis of X-linkage not only explained the original data, but it also predicted the results of other crosses. One important test is outlined in Fig. 17.5b. Here, the parental cross is the same as that in Fig. 17.5a, but instead of mating the F1 females to their brothers, they are mated to white-eyed males. The prediction is that there should be a 1:1 ratio of red-eyed to white-eyed females as well as a 1:1 ratio of red-eyed to white-eyed males. Again, these were the results observed. By the results of these crosses and others, Morgan demonstrated that the pattern of inheritance of the white-eye mutation parallels the pattern of inheritance of the X chromosome.