Simple multicellularity is widespread among eukaryotes.
A recent survey of eukaryotic organisms recognized 119 major groups within the superkingdoms discussed in Chapter 27. Of these, 83 contain only single-celled organisms, predominantly cells that engulf other microorganisms or ingest small organic particles, photosynthetic cells that live suspended in the water column, or parasitic cells that live within other organisms. Each of the 36 remaining branches exhibits some cases of simple multicellularity, mostly in the form of filaments, hollow balls, or sheets of little-differentiated cells (Fig. 28.1).
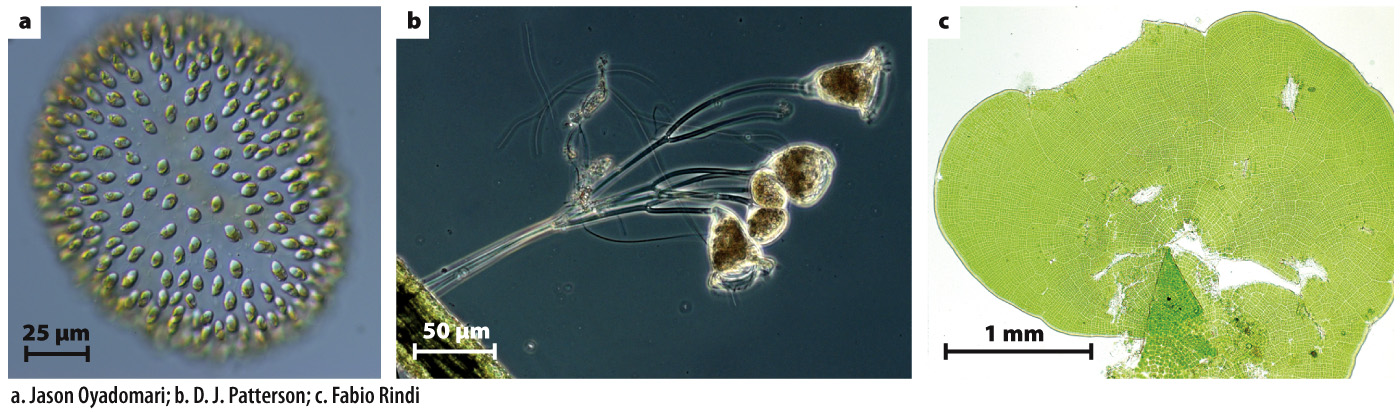
FIG. 28.1 Simple multicellularity. Cells in simple multicellular organisms show little differentiation and remain in close contact with the external environment. Many groups of eukaryotic organisms display simple multicellularity: (a) Uroglena, a stramenopile alga found commonly in lakes; (b) Epistylis, a stalked ciliate protozoan that lives attached to the surfaces of fish and crabs; and (c) Prasiola, a bladelike green alga only a single cell thick, found on tree trunks and rock surfaces.
Simple multicellular eukaryotes share several properties. In many, cell adhesion molecules cause adjacent cells to stick together, but there are few specialized cell types and relatively little communication or transfer of resources between cells. Most or all of the cells in simple multicellular organisms retain a full range of functions, including reproduction, and so the organism usually pays only a small penalty for individual cell death. Importantly, in simple multicellular organisms, nearly every cell is in direct contact with the external environment, at least during phases of the life cycle when the cells must acquire nutrients.
Simple multicellularity occurs most prominently among algae, although stalklike colonies of particle feeders have evolved in at least three groups of heterotrophic protists, and simple filamentous fungi absorb organic molecules as sources of carbon and energy. In addition, four eukaryotic groups have achieved simple multicellularity by a different route, aggregating during just one stage of the life cycle. Slime molds are the best-known example (Chapter 27).
Page 579
Six groups (two algal, three protozoan, and one fungal) include species characterized by coenocytic organization. In coenocytic organisms, the nucleus divides multiple times, but the cell does not, so the nuclei are not partitioned into individual cells. The result is a large cell—sometimes even visible to the naked eye—with many nuclei. The green algae Codium and Caulerpa, found along the shorelines of temperate and tropical seas, respectively, are common examples of coenocytic organisms (Fig. 28.2). Coenocytic rhizarians on the deep seafloor can be 10 to 20 cm long. There is no evidence that any coenocytic organisms evolved from truly multicellular ancestors, nor have any given rise to complex multicellular descendants.

FIG. 28.2 Coenocytic organization. Nuclei divide repeatedly but are not partitioned into individual cells. Two different groups of green algae have evolved large size as coenocytic organisms: (a) Codium, found abundantly along temperate coastlines, and (b) Caulerpa, common in shallow tropical seas and aquaria.
What selection pressures favored the evolution of simple multicellular organisms from single-celled ancestors? One selective advantage is that multicellularity helps organisms avoid getting eaten. In an illuminating experiment, single-celled green algae were grown in the presence of a protistan predator. Within 10 to 20 generations, most of the algae were living in eight-cell colonies that were essentially invulnerable to predator attack. Another advantage is that multicellular organisms may be able to maintain their position on a surface or in the water column better than their single-celled relatives. Seaweeds, for example, live anchored to the seafloor in places where light and nutrients support growth. Feeding provides a third potential advantage: In colonial heterotrophs such as the stalked ciliate Epistylis (see Fig. 28.1b), the coordinated beating of flagella assists feeding by directing currents of food-laden water toward the cells.
There are also costs associated with multicellularity, particularly for complex multicellular organisms with differentiated reproductive tissues. In these organisms, most cells do not reproduce, instead supporting the few that do. This requires cooperation among cells, but it creates opportunities for cells to “cheat”—to use nutrients for their own proliferation rather than the growth and reproduction of the organism as a whole. Cancer is the most conspicuous and lethal example of non-cooperation, perhaps the defining disease of complex multicellular organisms.
As we will see, complex multicellular organisms evolved from simple multicellular ancestors, but most taxonomic groups containing simple multicellular organisms never gave rise to complex descendants.