CAM plants use nocturnal CO2 storage to avoid water loss during the day.
In most plants, water loss and photosynthesis peak at the same time, when the sun is high overhead. What if a plant could open its stomata to capture CO2 at night, when cool air limits rates of evaporation?
In fact, a number of plants have evolved precisely this mechanism, called crassulacean acid metabolism, or CAM, to help balance CO2 gain and water loss (Fig. 29.6). CAM (named after the Crassulaceae family of plants, which uses this pathway) provides a system for storing CO2 overnight by converting it into a form that will not diffuse away. The storage form of CO2 is produced by the activity of the enzyme PEP carboxylase, which combines a dissolved form of CO2 (bicarbonate ion, HCO3–) with a 3-
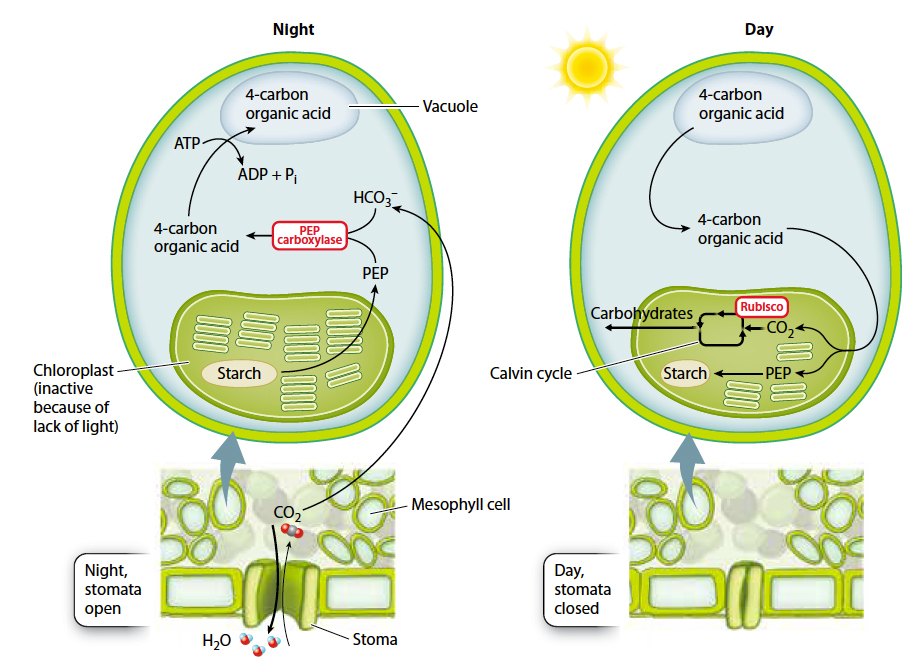
When the sun comes up the next morning, the stomata close, conserving water. At the same time, the 4-
By opening stomata only at night, CAM plants greatly increase the amount of CO2 gain per unit of water loss. Indeed, the CO2:H2O exchange ratio for CAM plants is in the range of 1:50, nearly 10 times higher than that of plants that open their stomata during the day. However, CAM has a drawback. The carbohydrate production by CAM photosynthesis is slower because ATP is needed to drive the uptake of organic acids into the vacuole and because only so much organic acid can accumulate overnight in the vacuole. CAM is therefore most common in habitats such as deserts, where water conservation is crucial, and among epiphytes, plants that grow on the branches of other plants, without contact with the soil.
Because CAM uses enzymes (for example, PEP carboxylase) and compartments (for example, vacuoles) that exist in all plant cells, it is not surprising that it has evolved multiple times. CAM occurs in all four groups of vascular plants shown in Fig. 29.1. It is most widespread in the angiosperms, occurring in 5% to 10% of species. Vanilla orchids and Spanish moss, two well-