Polar transport of auxin guides the placement of leaf primordia and the development of vascular connections with the stem.
The plant hormone auxin was first identified when it was observed to cause shoots to elongate. As we will see in section 31.5, auxin’s ability to trigger cell elongation underlies how shoots bend and grow toward light. Auxin also plays an important role in establishing spatial patterns of shoot development. In particular, auxin influences the spacing and placement of leaf primordia and the formation of the vascular bundles connecting young leaves to the mature vascular system of the stem.
Auxin’s role in patterning results from the fact that its direction of movement across a group of cells can be controlled. The net movement of auxin in a single direction creates regions of high and low concentrations of auxin. These gradients in auxin concentration can guide the growth and development of individual cells.
Let’s consider first how auxin movement influences the placement of new leaf primordia. Auxin is synthesized in rapidly dividing cells within the shoot meristem and is transported in the outermost cell layer toward the shoot tip. High levels of auxin in the outermost layer of cells trigger the growth of new leaf primordia. The movement of auxin into leaf primordia depletes auxin from surrounding regions and thus prevents new leaf primordia from forming nearby. As the shoot continues to grow, already formed leaf primordia become displaced from the shoot tip and spread farther and farther apart. This allows auxin moving upward in the outermost cell layer to accumulate to levels needed to trigger new leaf primordia close to the shoot tip.
Let’s now consider how auxin transport guides the formation of vascular bundles that connect the leaf to the stem. As auxin is transported into leaf primordia, it promotes cell expansion and the young leaves grow. However, auxin does not remain in the developing leaf. Instead, auxin that has accumulated at the tip of the young leaf is transported back toward the stem, but this time through cells in the interior of the leaf. As auxin moves through these cells, it causes the cells both to elongate and to produce more of the membrane transport proteins needed for auxin to exit the cell. The cells that initially, by chance, have the highest auxin levels become highly efficient at transporting auxin toward the stem. This, in turn, creates distinct “channels” through which auxin drains from the leaf. These continuous strands of elongate procambial cells eventually develop into veins and vascular bundles containing mature xylem and phloem.
These two examples illustrate how the movement of auxin in a single direction establishes spatial patterns that can guide further differentiation and development. But they leave open the question of how undifferentiated cells control the movement of auxin in the first place.
The coordinated movement of auxin across many cells in a single direction is referred to as polar transport (Fig. 31.9). Polar transport allows auxin to move independent of gravity at rates faster than the hormone could move by diffusion alone. However, it requires an input of energy.
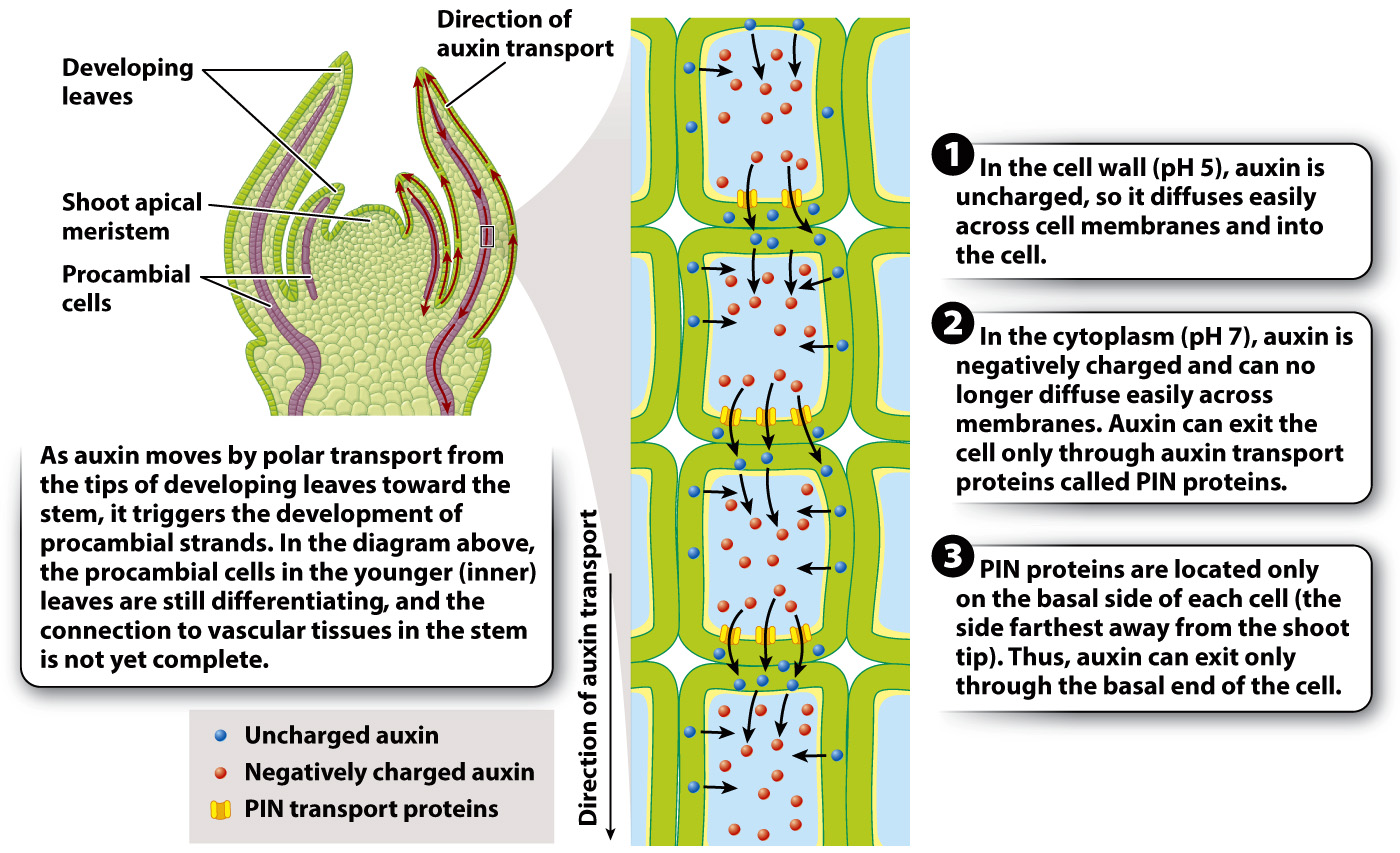
Polar transport depends on the difference in pH between the cell wall and the cytoplasm. In the cell wall, auxin carries no net charge and thus can enter surrounding cells by diffusing across the plasma membrane. However, because the pH of the cytoplasm is higher than the pH of the cell wall, auxin loses a proton. As a result, auxin has a net negative charge when inside the cell and cannot move easily across the plasma membrane. For auxin to leave the cell, an auxin-