Monocots are diverse in shape and size despite not forming a vascular cambium.
Monocots or monocotyledons make up nearly one-
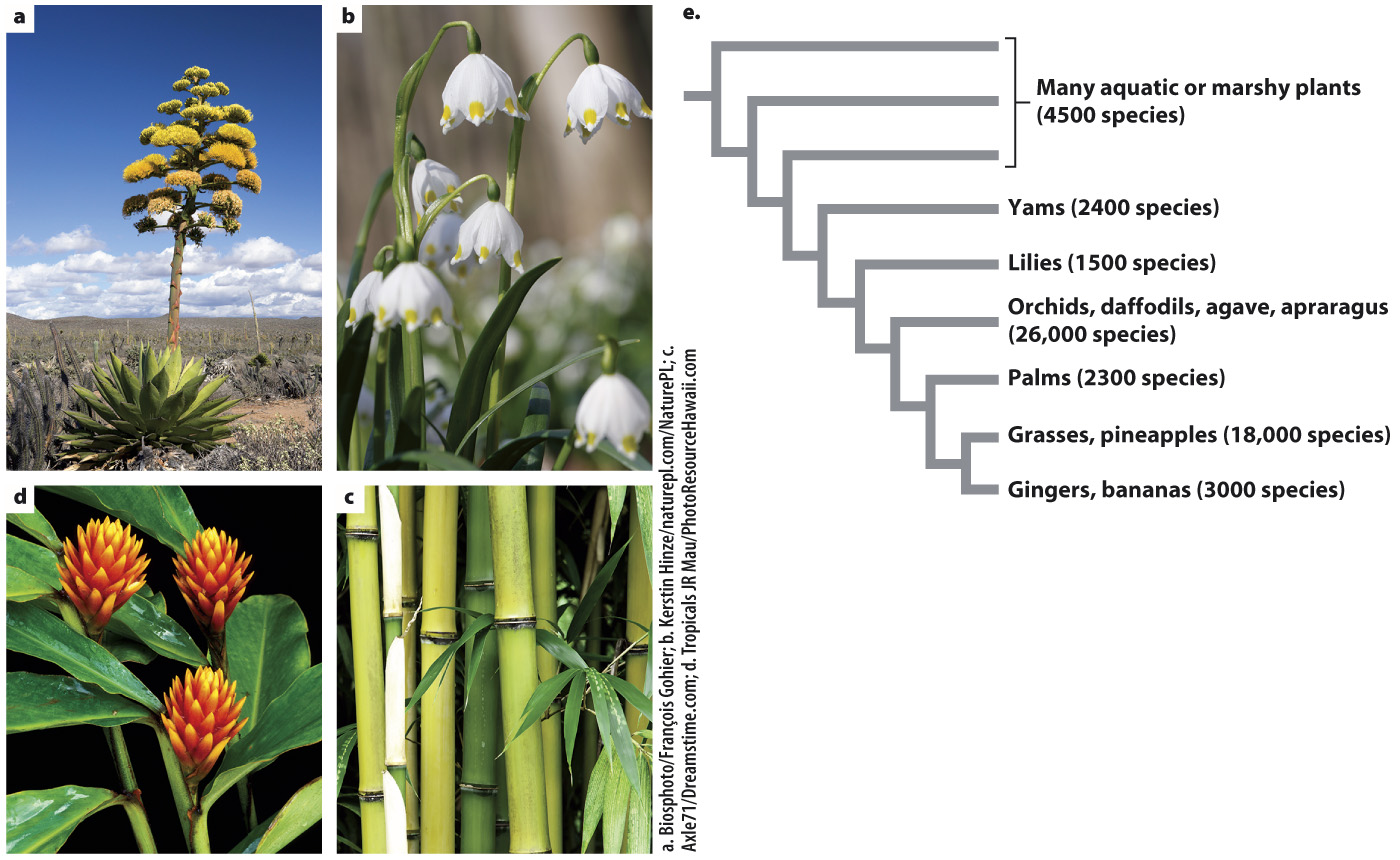
Monocots take their name from the fact that they have one embryonic seed leaf, or cotyledon, whereas all other angiosperms have two. However, monocots are distinct in form in so many other ways that one rarely has to count the number of cotyledons to identify a member of this large, monophyletic group. Monocots represent a major evolutionary departure in the way seed plants build their bodies, in that a vascular cambium is never formed. How then has this group been so successful?
In monocot leaves, the major veins are typically parallel and the base of the leaf surrounds the stem, forming a continuous sheath. This type of leaf base means that only one leaf can be attached at any node, consistent with the formation of a single cotyledon. Although monocots do not form a vascular cambium, they can still produce stems that are quite large. For example, both corn and coconut palms are monocots. In monocots, all of the increase in stem diameter occurs in a narrow zone immediately below the apical meristem. As a result of this lateral expansion, the vascular bundles become distributed throughout the stem instead of being arranged in a ring as in all other seed plants. Monocots that have slender stems—
The lack of a vascular cambium has a profound impact on the way monocots form roots. Because individual roots cannot increase their vascular capacity, monocots continuously initiate new roots from their stems. Thus, the root systems of monocots are more similar to those found in ferns and lycophytes than in other seed plants. Finally, monocot flowers typically produce organs in multiples of 3 (for example, having 3, 6, or 9 stamens), whereas eudicot flowers typically produce organs in multiples of 4 or 5.
Monocots have a relatively poor fossil record, in part because they do not produce wood. What factors might have led to such a radical revision in how they build their bodies? This question requires that we look at where and how living monocots grow. One hypothesis is that monocots evolved from ancestors that produced creeping, horizontal stems as they grew along the shores of lakes and other wetlands. Many monocots today, including the earliest diverging groups of monocots, grow in such habitats, and many of the features of the monocot body plan are well adapted to environments with loose substrates, flowing water, and fluctuating water levels. For example, their leaf base provides a firm attachment that prevents leaves from being pulled off by flowing water. Furthermore, many monocots produce strap-
We may never know for sure in what environment the monocots first evolved. We thus turn our attention to one of the most diverse groups within the monocots: the grasses. Many grasses produce stems that grow horizontally and branch, allowing them to cover large areas. Grasses have linear leaves that elongate from the base, allowing them to survive grazing as well as fire and drought. Many grasses have evolved the ability to tolerate dry environments by producing roots that extend deep into the soil. In addition, C4 photosynthesis has evolved within the grasses multiple times. As described in Chapter 29, C4 photosynthesis allows plants to avoid photorespiration and thus photosynthesize with greater efficiency. Grasses are among the most successful group of plants, becoming widespread within the past 20 million years as climates changed (Fig. 33.22). Today, nearly 30% of terrestrial environments are grasslands.
HOW DO WE KNOW?
FIG. 33.22
When did grasslands expand over the land surface?
BACKGROUND Today, prairies, steppes, and other grasslands occur widely in the interiors of continents. Grasses, however, do not fossilize readily. How, then, can we understand how grasslands developed through time?
HYPOTHESIS Grasslands expanded as climate changed over the past 50 million years.
OBSERVATIONS AND EXPERIMENTS Grasses commonly make phytoliths, small structures of silica (SiO2) in their cells. These preserve well, providing a direct record of grass expansion. Moreover, mammals that feed on grasses evolved high-
RESULTS Studies of phytoliths, mammal tooth structure, and carbon isotopic composition of teeth enamel from the North American midcontinent clearly show that grasslands expanded 20 to 15 million years ago, and C4 grasslands expanded later, about 8 to 6 million years ago.
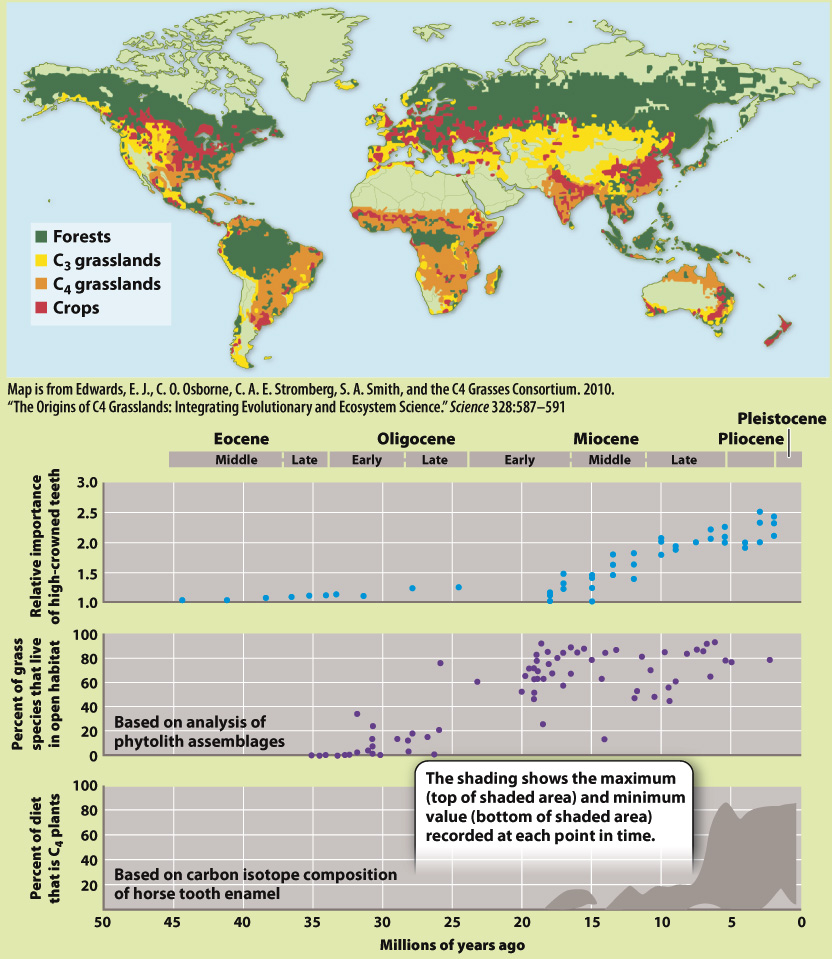
CONCLUSION In North America, grasslands expanded as atmospheric CO2 levels declined and climates became drier. Other continents show evidence of a similar linkage of grassland expansion to climate change.
FOLLOW-
SOURCE Stömberg, C. A. E. 2011. “Evolution of Grasses and Grassland Ecosystems.” Annual Review of Earth and Planetary Sciences 39: 517–
Quick Check 4 What are some of the distinctive features of monocots?
Quick Check 4 Answer
Monocots produce only a single cotyledon; monocot roots are produced continuously directly from the stem; many monocots produce creeping stems; and the strap-