Cellular DNA molecules take the form of a double helix.
To the knowledge of the chemical makeup of the nucleotides and their linkages in a DNA strand, Watson and Crick added results from earlier physical studies indicating that DNA is a long molecule. They also relied on important information from X-
With these critical pieces of information in hand, Watson and Crick set out to build a model of DNA that could account for the results of all previous chemical and physical experiments, using sheet metal cutouts of the bases and wire ties for the sugar–
Why all the fuss (and why the Nobel Prize nine years later)? First, let’s look at the structure, and you will see that the structure itself tells you how DNA carries and transmits genetic information. The Watson–
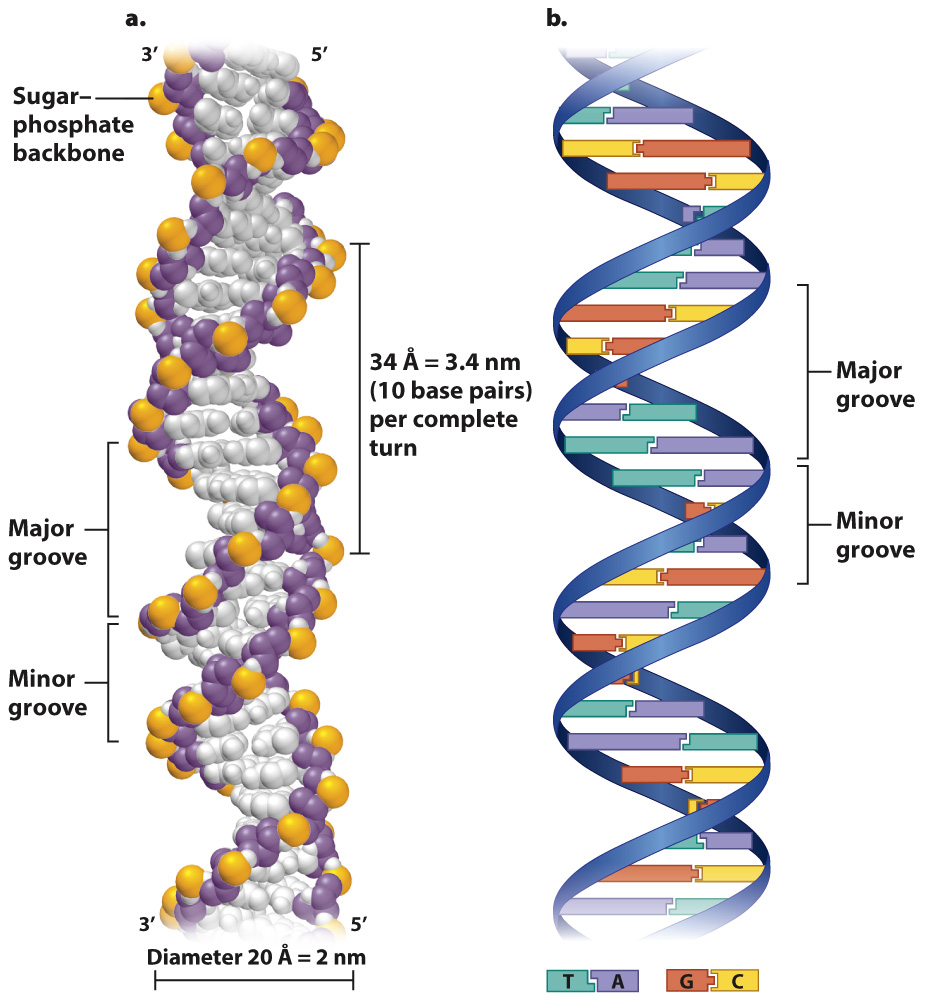
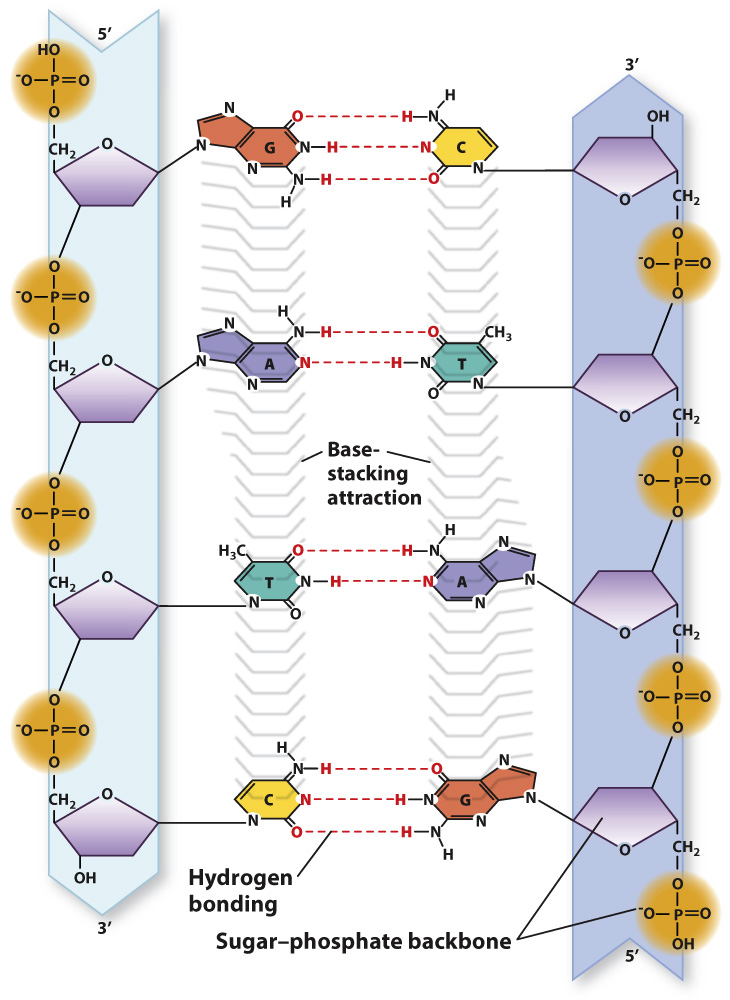
Importantly, the individual DNA strands in the double helix are antiparallel, which means that they run in opposite directions. That is, the 3′ end of one strand is opposite the 5′ end of the other. In Fig. 3.8a, the strand that starts at the bottom left and coils upward begins with the 3′ end and terminates at the top with the 5′ end, whereas its partner strand begins with its 5′ end at the bottom and terminates with the 3′ end at the top.
Fig. 3.8b shows a different depiction of double-
Note that, as shown in Fig. 3.8b, an A in one strand pairs only with a T in the other, and G pairs only with C. Each base pair contains a purine and a pyrimidine. This precise pairing maintains the structure of the double helix since pairing two purines would cause the backbones to bulge and pairing two pyrimidines would cause them to narrow. The pairing of one purine with one pyrimidine preserves the distance between the backbones along the length of the entire molecule.
Because they form specific pairs, the bases A and T are said to be complementary, as are the bases G and C. The formation of only A–
5′-ATGC-
3′-TACG-
Where one strand has the base A, the other strand across the way has the base T, and where one strand has a G, the other has a C. In other words, the paired strands are not identical but complementary. Because of the A–
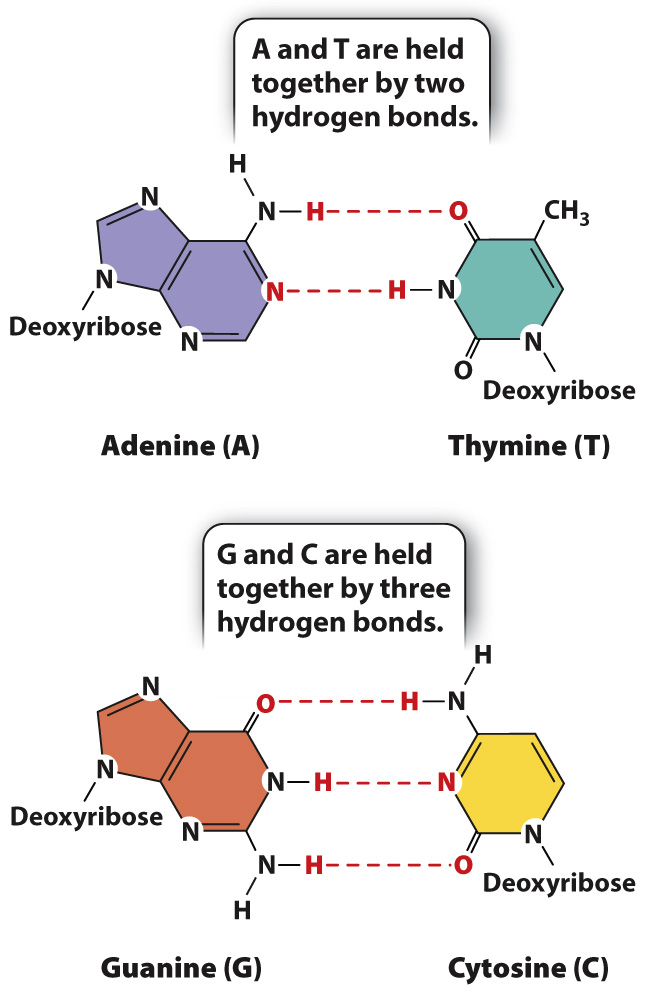
Why is it that A pairs only with T, and G only with C? Fig. 3.9 illustrates the answer. The specificity of base pairing is brought about by hydrogen bonds that form between A and T (two hydrogen bonds) and between G and C (three hydrogen bonds). A hydrogen bond in DNA is formed when an electronegative atom (O or N) in one base shares a hydrogen atom (H) with another electronegative atom in the base across the way. Hydrogen bonds are relatively weak bonds, typically 5% to 10% of the strength of covalent bonds, and can be disrupted by high pH or heat. However, in total they contribute to the stability of the DNA double helix.
An almost equally important factor contributing to the stability of the double helix is the interactions between bases in the same strand (Fig. 3.10). This stabilizing force is known as base stacking, and it occurs because the nonpolar, flat surfaces of the bases tend to group together away from water molecules, and hence stack on top of one another as tightly as possible.