Biology, Cognition, and Learning
 From drooling dogs, running rats, and pecking pigeons, we have learned much about the basic processes of learning. But conditioning principles don’t tell us the whole story. Once again we see one of psychology’s big ideas at work. Our learning is the product of the interaction of biological, psychological, and social-cultural influences.
From drooling dogs, running rats, and pecking pigeons, we have learned much about the basic processes of learning. But conditioning principles don’t tell us the whole story. Once again we see one of psychology’s big ideas at work. Our learning is the product of the interaction of biological, psychological, and social-cultural influences.
Biological Limits on Conditioning
6-11 What limits does biology place on conditioning?
Evolutionary theorist Charles Darwin proposed that natural selection favors traits that aid survival. In the middle of the twentieth century, researchers further showed that there are biological constraints on learning. Each species comes predisposed (biologically prepared) to learn those things crucial to its survival.
Limits on Classical Conditioning

A discovery by John Garcia and Robert Koelling in the 1960s helped end a popular and widely held belief in psychology: that environments rule our behavior. Part of this idea was that almost any stimulus (whether a taste, sight, or sound) could serve equally well as a conditioned stimulus. Garcia and Koelling’s work put that idea to the test and proved it wrong. They noticed that rats would avoid a taste—but not sights or sounds—associated with becoming sick, even hours later (1966). This response, which psychologists call taste aversion, makes adaptive sense. For rats, the easiest way to identify tainted food is to taste it. Taste aversion makes it tough to wipe out an invasion of “bait-shy” rats by poisoning. After being sickened by the bait, they are biologically prepared to avoid that taste ever after.
Humans, too, seem biologically prepared to learn some things rather than others. If you become violently ill four hours after eating a tainted hamburger, you will probably develop an aversion to the taste of hamburger. But you usually won’t avoid the sight of the associated restaurant, its plates, the people you were with, or the music you heard there.
Though Garcia and Koelling’s taste-aversion research began with the discomfort of some laboratory animals, it later enhanced the welfare of many others. In one taste-aversion study, coyotes and wolves were tempted into eating sheep carcasses laced with a sickening poison. Ever after, they avoided sheep meat (Gustavson et al., 1974, 1976). Two wolves penned with a live sheep seemed actually to fear it. These studies not only saved the sheep from their predators, but also saved the sheep-shunning coyotes and wolves from angry ranchers and farmers. In later experiments, conditioned taste aversion has successfully prevented baboons from raiding African gardens, raccoons from attacking chickens, ravens and crows from feeding on crane eggs, and Mexican wolves from preying on sheep. In all these cases, research helped preserve both the prey and their predators (Dingfelder, 2010; Garcia & Gustavson, 1997).

Such research supports Darwin’s principle that natural selection favors traits that aid survival. Our ancestors who readily learned taste aversions were unlikely to eat the same toxic food again and were more likely to survive and leave descendants. Nausea, like anxiety, pain, and other bad feelings, serves a good purpose. Like a car’s low-oil warning light, each alerts the body to a threat (Neese, 1991).
This tendency to learn behaviors favored by natural selection may help explain why we humans seem naturally disposed to learn associations between the color red and sexuality. Female primates display red when nearing ovulation. In human females, enhanced bloodflow produces the red blush of flirtation and sexual excitation. Does the frequent pairing of red and sex—with Valentine’s hearts, red-light districts, and red lipstick—naturally enhance men’s attraction to women? Experiments (FIGURE 6.8) indicate that it does (Elliot et al., 2013; Pazda & Elliot, 2012). 

RETRIEVE + REMEMBER
Question 6.13
 How did Garcia and Koelling’s taste-aversion studies help disprove the belief that almost any stimulus (tastes, sights, sounds) could serve equally well as a conditioned stimulus? Explain.
How did Garcia and Koelling’s taste-aversion studies help disprove the belief that almost any stimulus (tastes, sights, sounds) could serve equally well as a conditioned stimulus? Explain.
Garcia and Koelling demonstrated that rats may learn an aversion to tastes, on which their survival depends, but not to sights or sounds.
Limits on Operant Conditioning

As with classical conditioning, nature sets limits on each species’ capacity for operant conditioning. Author and humorist Mark Twain (1835–1910) said it well: “Never try to teach a pig to sing. It wastes your time and annoys the pig.”
We most easily learn and retain behaviors that reflect our biological predispositions. Thus, using food as a reinforcer, you could easily condition a hamster to dig or to rear up, because these are among the animal’s natural food-searching behaviors. But you won’t be so successful if you use food to try to shape face washing and other hamster behaviors that normally have no link to food or hunger (Shettleworth, 1973). Similarly, you could easily teach pigeons to flap their wings to avoid being shocked, and to peck to obtain food. That’s because fleeing with their wings and eating with their beaks are natural pigeon behaviors. However, pigeons have a hard time learning to peck to avoid a shock, or to flap their wings to obtain food (Foree & LoLordo, 1973). The principle: Our biology predisposes us to learn associations that are naturally adaptive.
Cognitive Influences on Conditioning
6-12 How do cognitive processes affect classical and operant conditioning?
Cognition and Classical Conditioning

John B. Watson, of the “Little Albert” research discussed earlier in this chapter, was one of many psychologists who built on Ivan Pavlov’s work. The two researchers shared many beliefs. They rejected “mentalistic” concepts (such as consciousness) that referred to inner thoughts, feelings, and motives (Watson, 1913). They also maintained that the basic laws of learning are the same for all animals—whether dogs or humans. Thus, the science of psychology should study how organisms respond to stimuli in their environments, said Watson. “Its theoretical goal is the prediction and control of behavior.” This view, that psychology should be an objective science based on observable behavior, was called behaviorism, and it influenced North American psychology during the first half of the twentieth century.
Later research has shown that Pavlov’s and Watson’s views of learning underestimated two important sets of influences. The first, as we have seen, is the way that biological predispositions limit our learning. The second is the effect of our cognitive processes—our thoughts, perceptions, and expectations—on learning. For example, people being treated for alcohol use disorder may be given alcohol spiked with a nauseating drug. However, their awareness that the drug, not the alcohol, causes the nausea tends to weaken the association between drinking alcohol and feeling sick. In classical conditioning, it is (especially with humans) not simply the CS-US pairing, but also the thought that counts.
Cognition and Operant Conditioning
B. F. Skinner granted the biological underpinnings of behavior and the existence of private thought processes. Nevertheless, many psychologists criticized him for discounting cognition’s importance.
A mere eight days before dying of leukemia in 1990, Skinner stood before those of us attending the American Psychological Association convention. In this final address, he again rejected the growing belief that cognitive processes have a necessary place in the science of psychology and even in our understanding of conditioning. For Skinner, thoughts and emotions were behaviors that follow the same laws as other behaviors.
Nevertheless, the evidence of cognitive processes cannot be ignored. For example, rats exploring a maze, given no obvious rewards, seem to develop a cognitive map, a mental representation of the maze. In one study, when an experimenter placed food in the maze’s goal box, these roaming rats ran the maze as quickly as (and even faster than) other rats that had always been rewarded with food for reaching the goal. Like people sightseeing in a new town, the exploring rats seemingly experienced latent learning during their earlier tours. Their latent learning became evident only when they had some reason to demonstrate it.

The cognitive perspective shows the limits of rewards. Promising people a reward for a task they already enjoy can backfire. Excessive rewards can destroy intrinsic motivation—the desire to do something well, for its own sake. In experiments, rewarding children with toys or candy for reading shortens the time they spend reading (Marinak & Gambrell, 2008). It is as if they think, “If I have to be bribed into doing this, it must not be worth doing for its own sake.”
To sense the difference between intrinsic motivation and extrinsic motivation (behaving in certain ways to gain external rewards or to avoid threatened punishment), think about your experience in this course. Are you feeling pressured to finish this reading before a deadline? Worried about your grade? Eager for the credits that will count toward graduation? If Yes, then you are extrinsically motivated (as, to some extent, almost all students must be). Are you also finding the material interesting? Does learning it make you feel more competent? If there were no grade at stake, might you be curious enough to want to learn the material for its own sake? If Yes, intrinsic motivation also fuels your efforts.
Nevertheless, rewards that signal a job well done—rather than to bribe or to control someone—can be effective (Boggiano et al., 1985). “Most improved player” awards, for example, can boost feelings of competence and increase enjoyment of a sport. Rightly administered, rewards can raise performance and spark creativity (Eisenberger & Rhoades, 2001; Henderlong & Lepper, 2002). And extrinsic rewards—such as the admissions, scholarships, and jobs that often follow hard work and good grades—are here to stay.
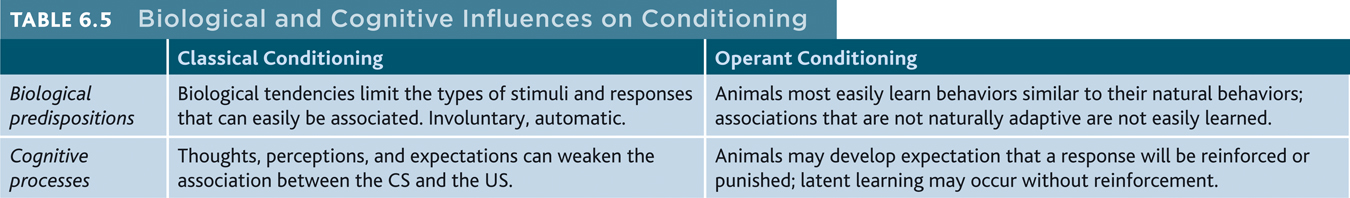
To sum up, TABLE 6.5 compares the biological and cognitive influences on classical and operant conditioning.
RETRIEVE + REMEMBER
Question 6.14
Latent learning is an example of what important idea?
The success of operant conditioning is affected not just by environmental cues, but also by cognitive factors.