The Plant Arabidopsis thaliana
Much of the early work in genetics was carried out on plants, including Mendel’s seminal discoveries in pea plants as well as important aspects of heredity, gene mapping, chromosome genetics, and quantitative inheritance in corn, wheat, beans, and other plants. However, by the mid-twentieth century, many geneticists had turned to bacteria, viruses, yeast, Drosophila, and mouse genetic models. Because a good genetic plant model did not exist, plants were relatively neglected, particularly for the study of molecular genetic processes.
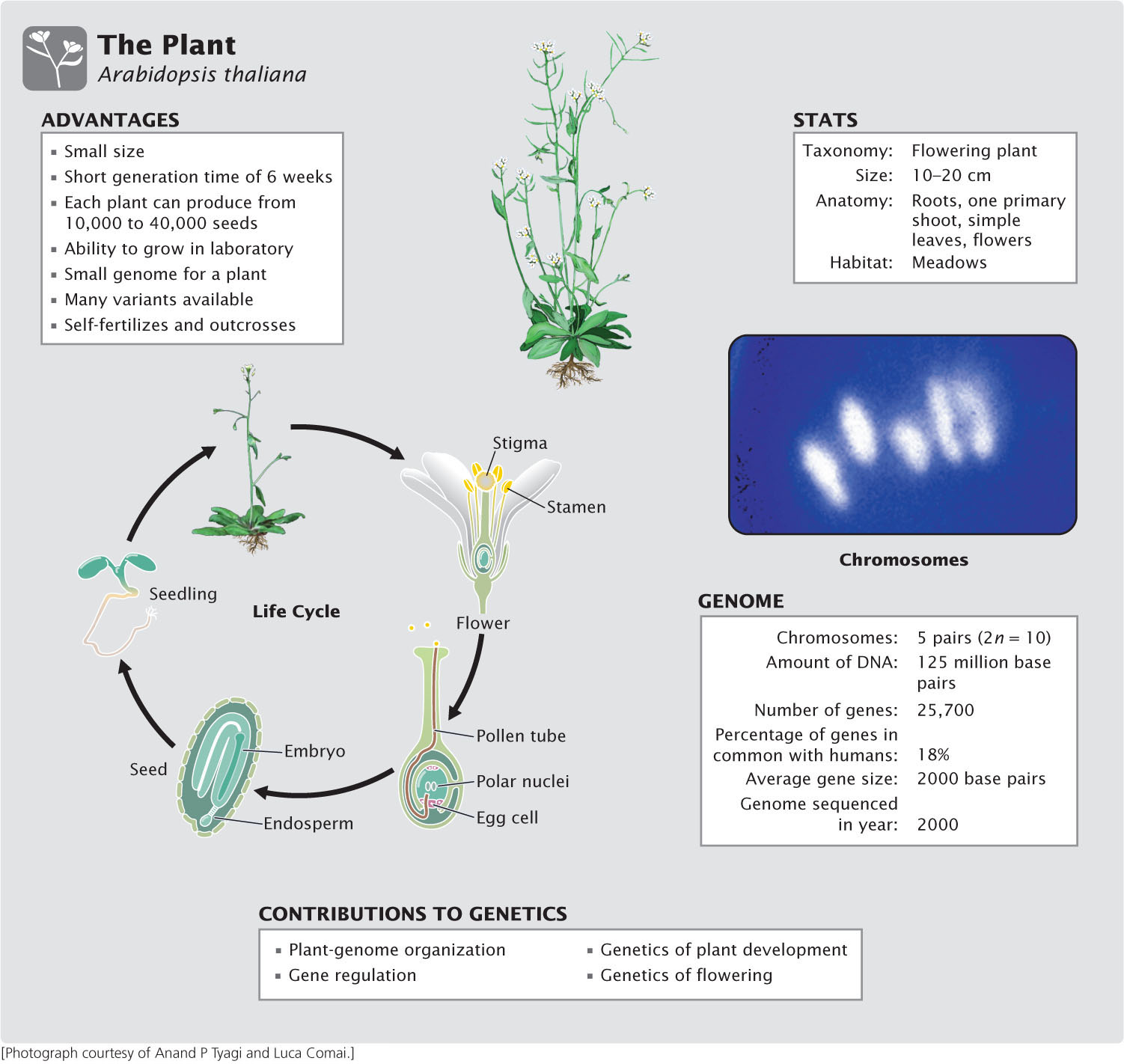
This changed in the last part of the twentieth century with the widespread introduction of a new genetic model organism, the plant Arabidopsis thaliana. A. thaliana was identified in the sixteenth century, and the first mutant was reported in 1873; but this species was not commonly studied until the first detailed genetic maps appeared in the early 1980s. Today, Arabidopsis figures prominently in the study of genome structure, gene regulation, development, and evolution in plants, and it provides important basic information about plant genetics that is applied to economically important plant species.
Advantages of Arabidopsis as a model genetic organism
The thale cress Arabidopsis thaliana is a member of the Brassicaceae family and grows as a weed in many parts of the world. Except in its role as a model genetic organism, Arabidopsis has no economic importance, but it has a number of characteristics that make it well suited to the study of genetics. As an angiosperm, it has features in common with other flowering plants, some of which play critical roles in the ecosystem or are important sources of food, fiber, building materials, and pharmaceutical agents. Arabidopsis’s chief advantages are its small size (maximum height of 10-20 cm), prolific reproduction, and small genome.
Arabidopsis thaliana completes development—from seed germination to seed production—in about 6 weeks. Its small size and ability to grow under low illumination make it ideal for laboratory culture. Each plant is capable of producing from 10,000 to 40,000 seeds, and the seeds typically have a high rate of germination; so large numbers of progeny can be obtained from single genetic crosses.
The Arabidopsis genome
A key advantage for molecular studies is Arabidopsis’s small genome, which consists of only 125 million base pairs of DNA on five pairs of chromosomes, compared with 2.5 billion base pairs of DNA in the maize genome and 16 billion base pairs in the wheat genome. The genome of A. thaliana was completely sequenced in 2000, providing detailed information about gene structure and organization in this species. A number of variants of A. thaliana—called ecotypes—that vary in shape, size, physiological characteristics, and DNA sequence are available for study.
Life cycle of Arabidopsis
The Arabidopsis life cycle is fairly typical of most flowering plants (see Figure 2.22). The main, vegetative part of the plant is diploid; haploid gametes are produced in the pollen and ovaries. When a pollen grain lands on the stigma of a flower, a pollen tube grows into the pistil and ovary. Two haploid sperm nuclei contained in each pollen grain travel down the pollen tube and enter the embryo sac. There, one of the haploid sperm cells fertilizes the haploid egg cell to produce a diploid zygote. The other haploid sperm cell fuses with two haploid nuclei to form the 3n endosperm, which provides tissue that will nourish the growing embryonic plant. The zygotes develop within the seeds, which are produced in a long pod.
Under appropriate conditions, the embryo germinates and begins to grow into a plant. The shoot grows upward and the roots downward, a compact rosette of leaves is produced and, under the right conditions, the shoot enlarges and differentiates into flower structures. At maturity, A. thaliana is a low-growing plant with roots, a main shoot with branches that bear mature leaves, and small white flowers at the tips of the branches.
Genetic techniques with Arabidopsis
A number of traditional and modern molecular techniques are commonly used with Arabidopsis and provide it with special advantages for genetic studies. Arabidopsis can self-fertilize, which means that any recessive mutation appearing in the germ line can be recovered in the immediate progeny. Cross-fertilization also is possible by removing the anther from one plant and dusting pollen on the stigma of another plant—essentially the same technique used by Gregor Mendel with pea plants (see Figure 3.4).
As already mentioned, many naturally occurring variants of Arabidopsis are available for study, and new mutations can be produced by exposing its seeds to chemical mutagens, radiation, or transposable elements that randomly insert into genes. The large number of offspring produced by Arabidopsis facilitates screening for rare mutations.
Genes from other organisms can be transferred to Arabidopsis by the Ti plasmid from the bacterium Agrobacterium tumefaciens, which naturally infects plants and transfers the Ti plasmid to plant cells (see Chapter 19). After transfer to a plant cell, the Ti plasmid randomly inserts into the DNA of the plant that it infects, thereby generating mutations in the plant DNA in a process called insertional mutagenesis. Geneticists have modified the Ti plasmid to carry a GUS gene, which has no promoter of its own. The GUS gene encodes an enzyme that converts a colorless compound (X-Glu) into a blue dye. Because the GUS gene has no promoter, it is expressed only when inserted into the coding sequence of a plant gene. When that happens, the enzyme encoded by GUS is synthesized and converts X-Glu into a blue dye that stains the cell. This dye provides a means to visually determine the expression pattern of a gene that has been interrupted by Ti DNA, producing information about the expression of genes that are mutated by insertional mutagenesis.
831
832