12.5 Recombination Takes Place Through the Breakage, Alignment, and Repair of DNA Strands
Recombination is the exchange of genetic information between DNA molecules; when the exchange is between homologous DNA molecules, it is called homologous recombination. This process takes place in crossing over, in which homologous regions of chromosomes are exchanged (see Figure 7.5) and alleles are shuffled into new combinations. Recombination is an extremely important genetic process because it increases genetic variation. Rates of recombination provide important information about linkage relations among genes, which is used to create genetic maps (see Figures 7.13 and 7.14). Recombination is also essential for some types of DNA repair (as will be discussed in Chapter 18).
Homologous recombination is a remarkable process: a nucleotide strand of one chromosome aligns precisely with a nucleotide strand of the homologous chromosome, breaks arise in corresponding regions of different DNA molecules, parts of the molecules precisely change place, and then the pieces are correctly joined. In this complicated series of events, no genetic information is lost or gained. Although the precise molecular mechanism of homologous recombination is still not well understood, the exchange is probably accomplished through the pairing of complementary bases. A single-stranded DNA molecule of one chromosome pairs with a single-stranded DNA molecule of another, forming heteroduplex DNA.
In meiosis, homologous recombination (crossing over) could theoretically take place before, during, or after DNA synthesis. Cytological, biochemical, and genetic evidence indicates that it takes place in prophase I of meiosis, whereas DNA replication takes place earlier, in interphase. Thus, crossing over must entail the breaking and rejoining of chromatids when homologous chromosomes are at the four-strand stage (see Figure 7.5). This section explores some theories about how the process of recombination takes place.
Models of Recombination
Homologous recombination can take place through several different pathways. One pathway is initiated by a single-strand break in each of two DNA molecules and includes the formation of a special structure called the Holliday junction (Figure 12.20). In this model, double-stranded DNA molecules from two homologous chromosomes align precisely. A single-strand break in one of the DNA molecules provides a free end that invades and joins the free end of the other DNA molecule. Strand invasion and joining take place on both DNA molecules, creating two heteroduplex DNAs, each consisting of one original strand plus one new strand from the other DNA molecule. The point at which nucleotide strands pass from one DNA molecule to the other is the Holliday junction. The junction moves along the molecules in a process called branch migration. The exchange of nucleotide strands and branch migration produce a structure termed the Holliday intermediate, which can be cleaved in one of two ways. Cleavage in the horizontal plane, followed by rejoining of the strands, produces noncrossover recombinants, in which the genes on either end of the molecules are identical with those originally present (gene A with gene B, and gene a with gene b). Cleavage in the vertical plane, followed by rejoining, produces crossover recombinants, in which the genes on either end of the molecules are different from those originally present (gene A with gene b, and gene a with gene B).
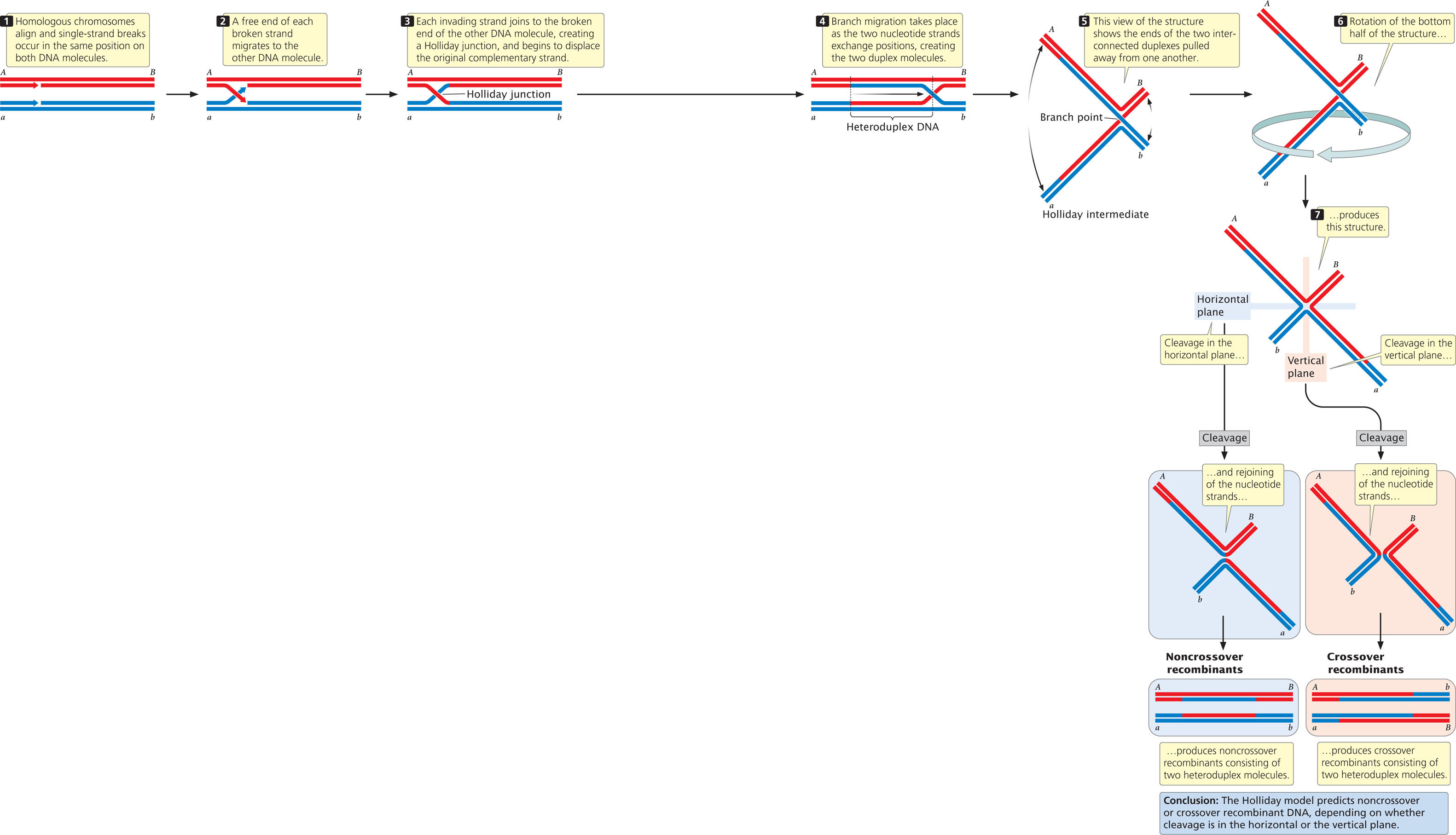
Another pathway for recombination is initiated by double-strand breaks in one of the two aligned DNA molecules (Figure 12.21). In this model, the removal of some nucleotides at the ends of the broken strands—followed by strand invasion, displacement, and replication—produces two heteroduplex DNA molecules joined by two Holliday junctions. The interconnected molecules produced in the double-strand-break model can be separated by further cleavage and reunion of the nucleotide strands in the same way that the Holliday intermediate is separated in the single-strand-break model. Whether crossover or noncrossover molecules are produced depends on whether cleavage is in the vertical or the horizontal plane. View  Animation 12.5 to see how the Holliday and double-strand break models lead to recombination.
Animation 12.5 to see how the Holliday and double-strand break models lead to recombination.
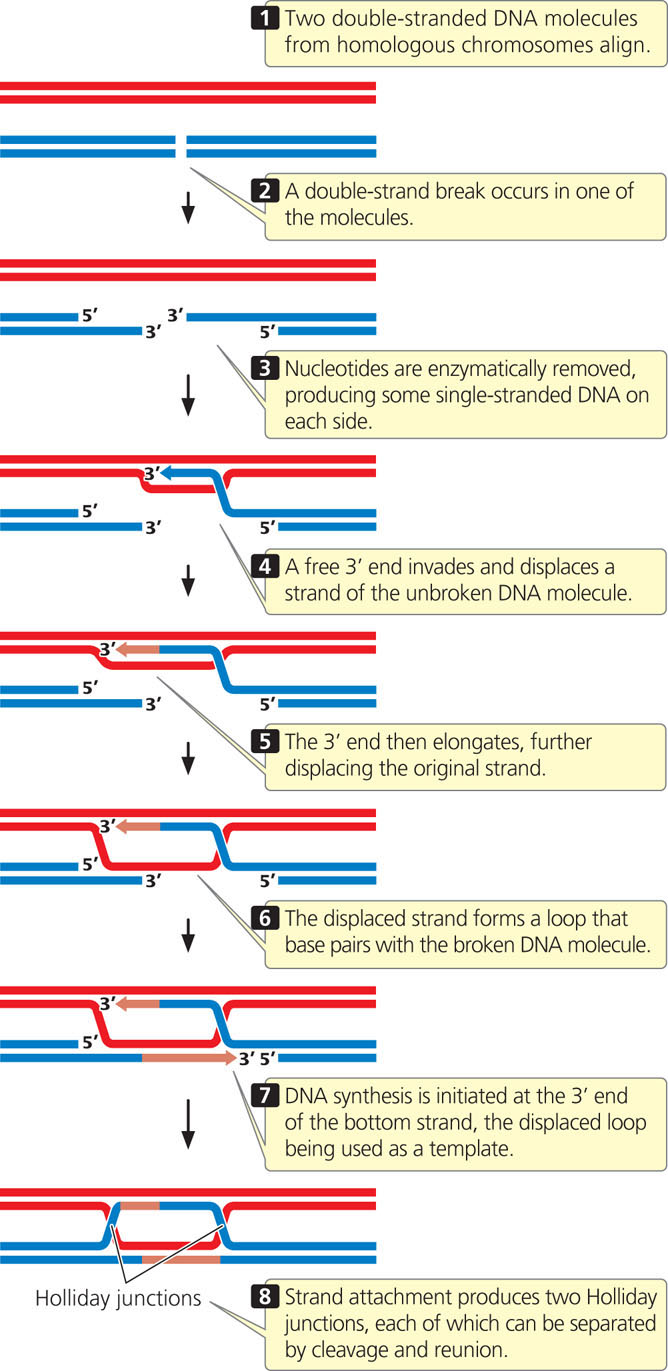
Evidence for the double-strand-break model originally came from results of genetic crosses in yeast that could not be explained by the Holliday model. Subsequent observations showed that double-strand breaks appear in yeast in prophase I, when crossing over takes place, and that mutant strains that are unable to form double-strand breaks do not exhibit meiotic recombination. Although considerable evidence supports the double-strand-break model in yeast, the extent to which it applies to other organisms is not yet known.
CONCEPTS
Homologous recombination requires the formation of heteroduplex DNA consisting of one nucleotide strand from each of two homologous chromosomes. In the Holliday model, homologous recombination is accomplished through a single-strand break in the DNA, strand displacement, and branch migration. In the double-strand-break model, recombination is accomplished through double-strand breaks, strand displacement, and branch migration.
 CONCEPT CHECK 11
CONCEPT CHECK 11Why is recombination important?
Enzymes Required for Recombination
Recombination between DNA molecules requires the unwinding of DNA helices, the cleavage of nucleotide strands, strand invasion, and branch migration, followed by further strand cleavage and union to remove Holliday junctions. Much of what we know about these processes arises from studies of gene exchange in E. coli. Although bacteria do not undergo meiosis, they do have a type of sexual reproduction (conjugation), in which one bacterium donates its chromosome to another (discussed more fully in Chapter 9). Subsequent to conjugation, the recipient bacterium has two chromosomes, which may undergo homologous recombination. Geneticists have isolated mutant strains of E. coli that are deficient in recombination; the study of these strains has resulted in the identification of genes and proteins that take part in bacterial recombination, revealing several different pathways by which it can take place.
Three genes that play pivotal roles in E. coli recombination are recB, recC, and recD, which encode three polypeptides that together form the RecBCD protein. This protein unwinds double-stranded DNA and is capable of cleaving nucleotide strands. The recA gene encodes the RecA protein; this protein allows a single strand to invade a DNA helix and the subsequent displacement of one of the original strands. In eukaryotes, the formation and branch migration of Holliday structures is facilitated by the enzyme Rad51.
In E. coli, ruvA and ruvB genes encode proteins that catalyze branch migration, and the ruvC gene produces a protein, called resolvase, that cleaves Holliday structures. Cleavage and resolution of Holliday structures in eukaryotes is carried out by an analogous enzyme called GEN1. Single-strand-binding proteins, DNA ligase, DNA polymerases, and DNA gyrase also play roles in various types of recombination, in addition to their functions in DNA replication.
CONCEPTS
A number of proteins have roles in recombination, including RecA, RecBCD, RuvA, RuvB, resolvase, single-strand-binding proteins, ligase, DNA polymerases, and gyrase.
CONCEPT CHECK 12What is the function of resolvase in recombination?
- Unwinds double-stranded DNA.
- Allows a single DNA strand to invade a DNA helix.
- Displaces one of the original DNA strands during branch migration.
- Cleaves the Holliday structure.
Gene Conversion
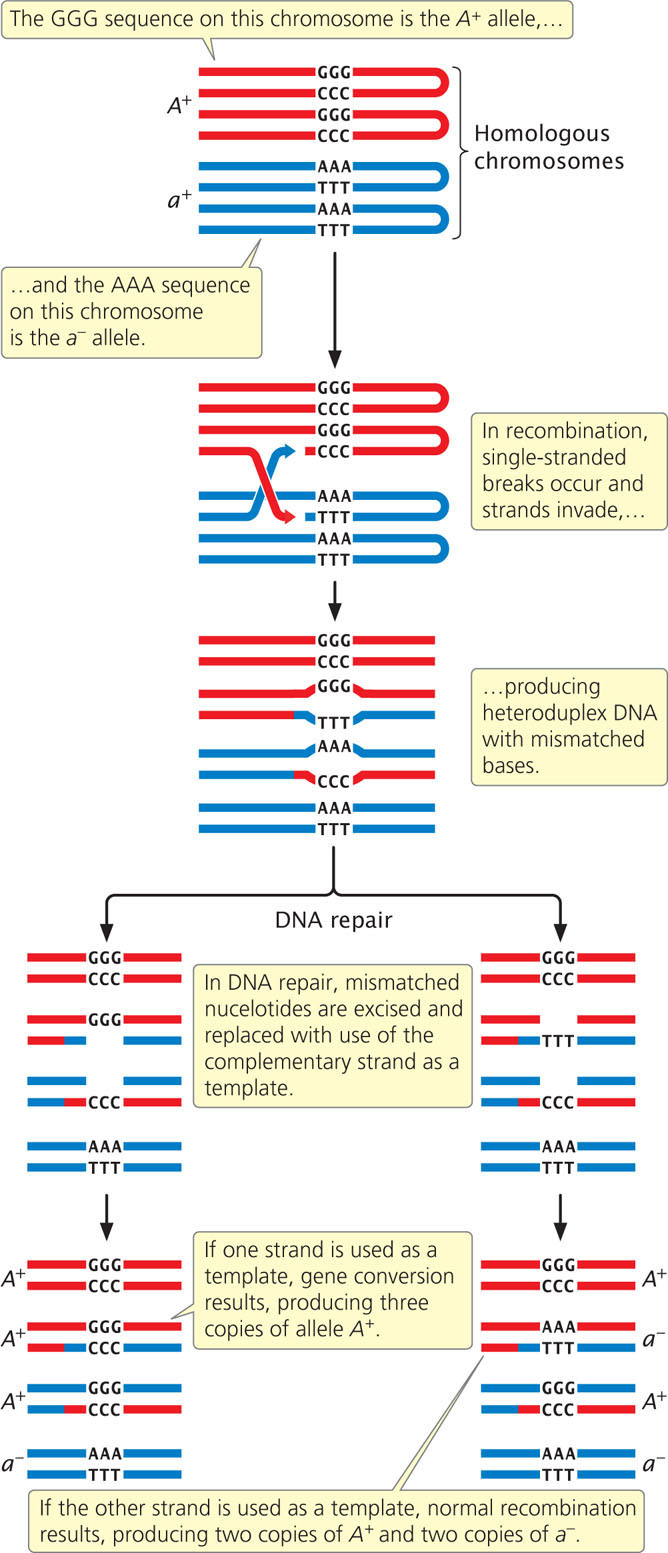
As we have seen, homologous recombination is the mechanism that produces crossing over. It is also responsible for a related phenomenon known as gene conversion, a process of nonreciprocal genetic exchange that can produce abnormal ratios of gametes following meiosis. For example, an individual organism with genotype Aa is expected to produce 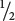 A gametes and
a gametes. Sometimes, however, meiosis in an Aa individual produces
A gametes and
a gametes. Sometimes, however, meiosis in an Aa individual produces 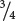 A and
A and 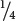 a or
A and
a. Gene conversion arises from heteroduplex formation that takes place in recombination. During heteroduplex formation, a single-stranded DNA molecule of one chromosome pairs with a single-stranded DNA molecule of another chromosome. If the two strands in a heteroduplex come from chromosomes with different alleles, there will be a mismatch of bases in the heteroduplex DNA (Figure 12.22). Such mismatches are often repaired by the cell. Repair mechanisms frequently excise nucleotides on one of the strands and replace them with new DNA by using the complementary strand as a template. One copy of an allele may be converted into the other allele, leading to a gene-conversion event (see Figure 12.22), depending on which strand serves as a template.
a or
A and
a. Gene conversion arises from heteroduplex formation that takes place in recombination. During heteroduplex formation, a single-stranded DNA molecule of one chromosome pairs with a single-stranded DNA molecule of another chromosome. If the two strands in a heteroduplex come from chromosomes with different alleles, there will be a mismatch of bases in the heteroduplex DNA (Figure 12.22). Such mismatches are often repaired by the cell. Repair mechanisms frequently excise nucleotides on one of the strands and replace them with new DNA by using the complementary strand as a template. One copy of an allele may be converted into the other allele, leading to a gene-conversion event (see Figure 12.22), depending on which strand serves as a template.