18.2 Mutations Are Potentially Caused by a Number of Different Factors
Mutations result from both internal and external factors. Those that occur under normal conditions are termed spontaneous mutations, whereas those that result from changes caused by environmental chemicals or radiation are induced mutations.
Spontaneous Replication Errors
Replication is amazingly accurate: less than one error in a billion nucleotides arises in the course of DNA synthesis (see Chapter 12). However, spontaneous replication errors do occasionally occur.
Tautomeric Shifts
The primary cause of spontaneous replication errors was formerly thought to be tautomeric shifts, in which the positions of protons in the DNA bases change. Purine and pyrimidine bases exist in different chemical forms called tautomers. The two tautomeric forms of each base are in dynamic equilibrium, although one form is more common than the other. The standard Watson-and-Crick base pairings—adenine with thymine, and cytosine with guanine—occur between the common forms of the bases, but, if the bases are in their rare tautomeric forms, other base pairings are possible. For example, the common form of cytosine pairs with guanine, but the rare tautomer of cytosine pairs with adenine.
Watson and Crick proposed that tautomeric shifts might produce mutations, and, for many years, their proposal was the accepted model for spontaneous replication errors. However, there has never been convincing evidence that the rare tautomers are the cause of spontaneous mutations. Furthermore, research now shows little evidence of tautomers in DNA.
504
Mispairing Due to Other Structures
Mispairings often arise through wobble (see Chapter 15), in which normal, protonated, and other forms of the bases are able to pair because of flexibility in the DNA helical structure (Figure 18.10). These structures have been detected in DNA molecules and are now thought to be responsible for many of the mispairings in replication.
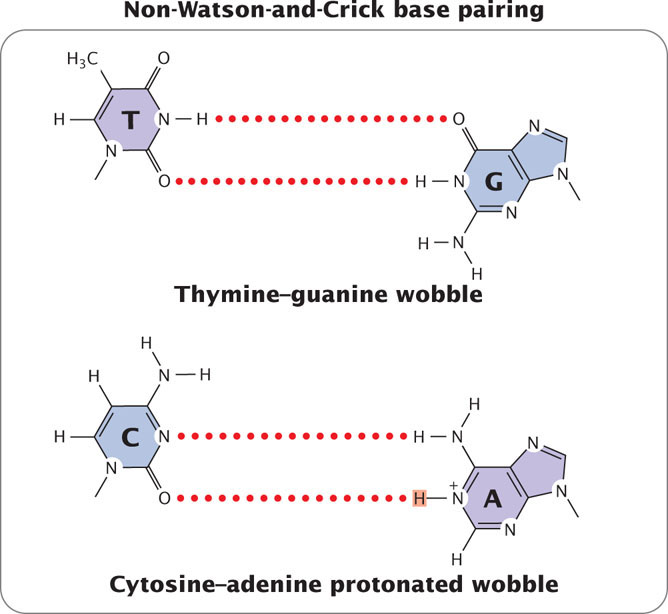
Incorporation Errors and Replicated Errors
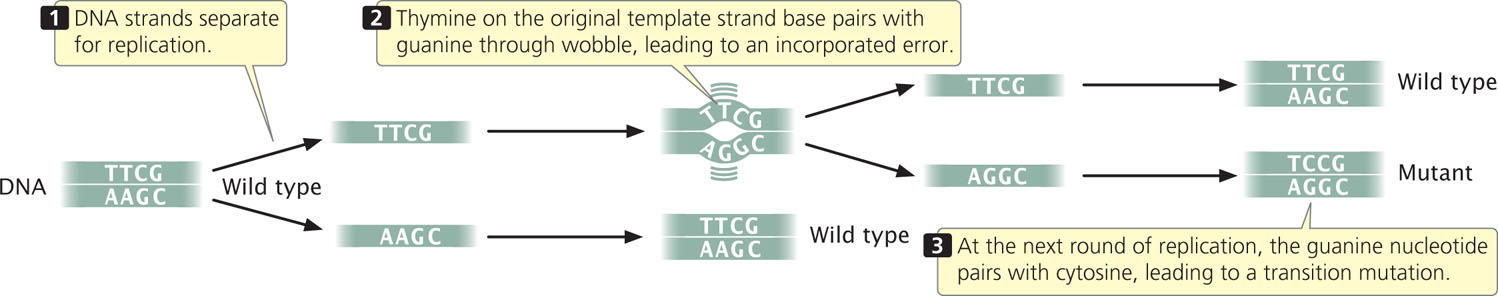
When a mismatched base has been incorporated into a newly synthesized nucleotide chain, an incorporated error is said to have occurred. Suppose that, in replication, thymine (which normally pairs with adenine) mispairs with guanine through wobble (Figure 18.11). In the next round of replication, the two mismatched bases separate, and each serves as template for the synthesis of a new nucleotide strand. This time, thymine pairs with adenine, producing another copy of the original DNA sequence. On the other strand, however, the incorrectly incorporated guanine serves as the template and pairs with cytosine, producing a new DNA molecule that has an error—a C · G pair in place of the original T · A pair (a T · A → C · G base substitution). The original incorporated error leads to a replicated error, which creates a permanent mutation because all the base pairings are correct and there is no mechanism for repair systems to detect the error.
Causes of Deletions and Insertions
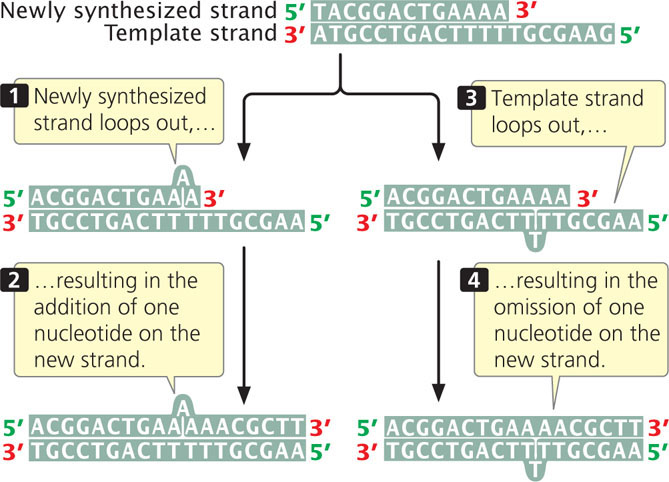
Mutations due to small insertions and deletions also arise spontaneously in replication and crossing over. Strand slippage can occur when one nucleotide strand forms a small loop (Figure 18.12). If the looped-out nucleotides are on the newly synthesized strand, an insertion results. At the next round of replication, the insertion will be replicated and both strands will contain the insertion. If the looped-out nucleotides are on the template strand, then the newly replicated strand will have a deletion, and this deletion will be perpetuated in subsequent rounds of replication.
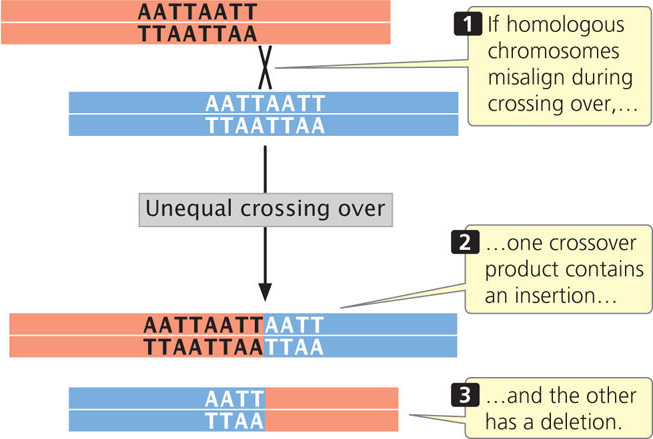
Another process that produces insertions and deletions is unequal crossing over. In normal crossing over, the homologous sequences of the two DNA molecules align, and crossing over produces no net change in the number of nucleotides in either molecule. Misaligned pairing can cause unequal crossing over, which results in one DNA molecule with an insertion and the other with a deletion (Figure 18.13).
CONCEPTS
Spontaneous replication errors arise from altered-base structures and from wobble-base pairing. Small insertions and deletions can occur through strand slippage in replication and through unequal crossing over.
Spontaneous Chemical Changes
In addition to spontaneous mutations that arise in replication, mutations also result from spontaneous chemical changes in DNA. One such change is depurination, the loss of a purine base from a nucleotide. Depurination results when the covalent bond connecting the purine to the 1′-carbon atom of the deoxyribose sugar breaks (Figure 18.14a), producing an apurinic site, a nucleotide that lacks its purine base. An apurinic site cannot act as a template for a complementary base in replication. In the absence of base-pairing constraints, an incorrect nucleotide (most often adenine) is incorporated into the newly synthesized DNA strand opposite the apurinic site (Figure 18.14b), frequently leading to an incorporated error. The incorporated error is then transformed into a replicated error at the next round of replication. Depurination is a common cause of spontaneous mutation; a mammalian cell in culture loses approximately 10,000 purines every day. Loss of pyrimidine bases also occurs, but at a much lower rate than depurination.
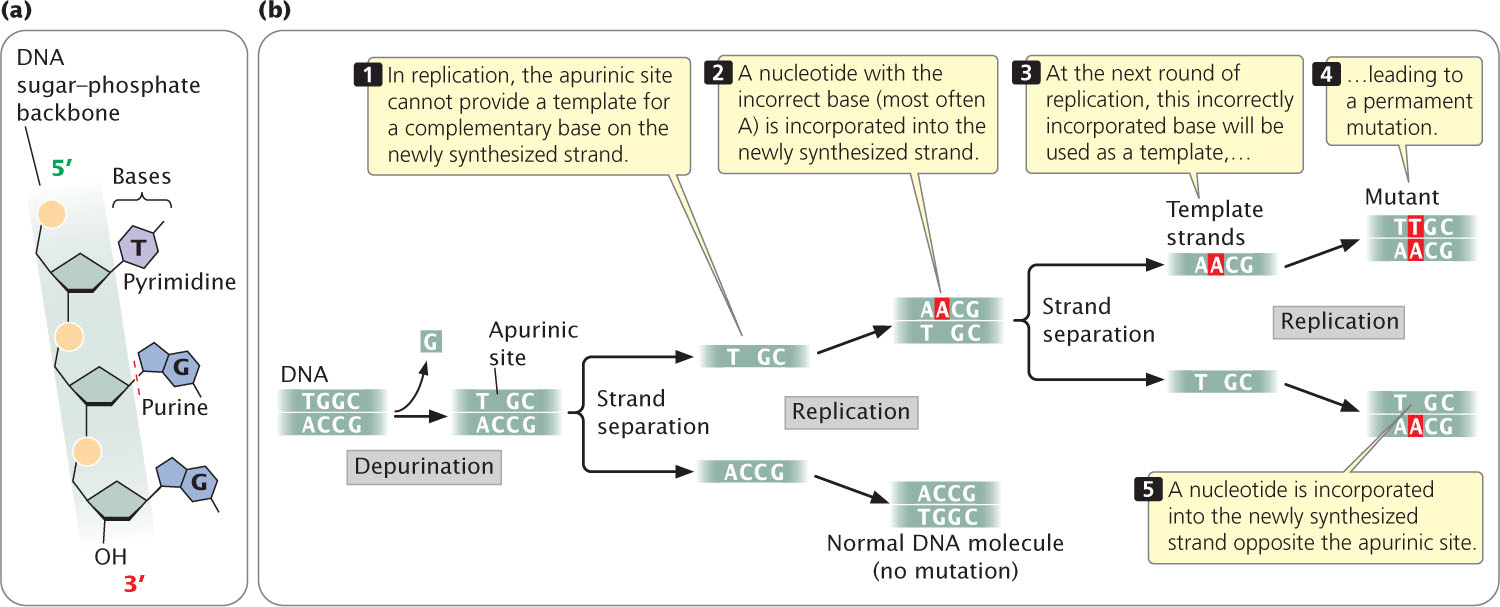
505
Another spontaneously occurring chemical change that takes place in DNA is deamination, the loss of an amino group (NH2) from a base. Deamination can be spontaneous or be induced by mutagenic chemicals.
Deamination can alter the pairing properties of a base: the deamination of cytosine, for example, produces uracil (Figure 18.15a), which pairs with adenine in replication. After another round of replication, the adenine will pair with thymine, creating a T · A pair in place of the original C · G pair (C · G → U · A → T · A); this chemical change is a transition mutation. This type of mutation is usually prevented by enzymes that remove uracil whenever it is found in DNA. The ability to recognize the product of cytosine deamination may explain why thymine, not uracil, is found in DNA. In mammals, including humans, some cytosine bases in DNA are naturally methylated and exist in the form of 5-methylcytosine (5mC; see Figure 10.19). When deaminated, 5mC becomes thymine (Figure 18.15b). Because thymine pairs with adenine in replication, the deamination of 5-methylcytosine changes an original C · G pair to T · A (C · G → 5mC · G → T · G → T · A). Consequently, C · G → T · A transitions are frequent in mammalian cells, and 5mC sites are mutation hotspots in humans. ![]() TRY PROBLEM 27
TRY PROBLEM 27
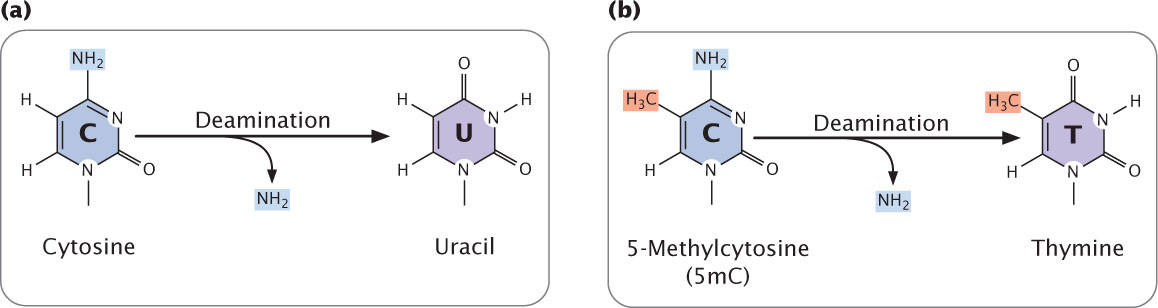
506
CONCEPTS
Some mutations arise from spontaneous alterations in DNA structure, such as depurination and deamination, which can alter the pairing properties of the bases and cause errors in subsequent rounds of replication.
Chemically Induced Mutations
Although many mutations arise spontaneously, a number of environmental agents are capable of damaging DNA, including certain chemicals and radiation. Any environmental agent that significantly increases the rate of mutation above the spontaneous rate is called a mutagen.
The first discovery of a chemical mutagen was made by Charlotte Auerbach, who started her career in Berlin researching the development of mutants in Drosophila. Faced with increasing anti-Semitism in Nazi Germany, Auerbach emigrated to Britain in 1933. There she continued her research on Drosophila and in 1940 began a collaboration with pharmacologist John Robson on the mutagenic effects of mustard gas, which had been used as a chemical weapon in World War I. The experimental conditions were crude. They heated liquid mustard gas over a Bunsen burner on the roof of the pharmacology building, and the flies were exposed to the gas in a large chamber. After developing serious burns on her hands from the gas, Auerbach let others carry out the exposures, and she analyzed the flies. Auerbach and Robson showed that mustard gas is indeed a powerful mutagen, reducing the viability of gametes and increasing the numbers of mutations seen in the offspring of exposed flies. Because the research was part of the secret war effort, publication of their findings was delayed until 1947.
Base Analogs
One class of chemical mutagens consists of base analogs, chemicals with structures similar to that of any of the four standard bases of DNA. DNA polymerases cannot distinguish these analogs from the standard bases; so, if base analogs are present during replication, they may be incorporated into newly synthesized DNA molecules. For example, 5-bromouracil (5BU) is an analog of thymine; it has the same structure as that of thymine except that it has a bromine (Br) atom on the 5-carbon atom instead of a methyl group (Figure 18.16a). Normally, 5-bromouracil pairs with adenine just as thymine does, but it occasionally mispairs with guanine (Figure 18.16b), leading to a transition (T · A → 5BU · A → 5BU · G → C · G), as shown in Figure 18.17. Through mispairing, 5-bromouracil can also be incorporated into a newly synthesized DNA strand opposite guanine. In the next round of replication 5-bromouracil pairs with adenine, leading to another transition (G · C → G · 5BU → A · 5BU → A · T).
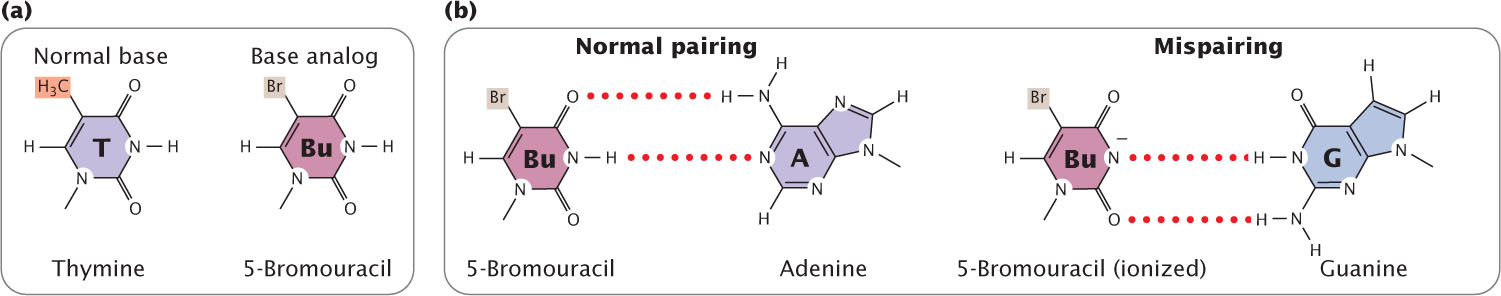
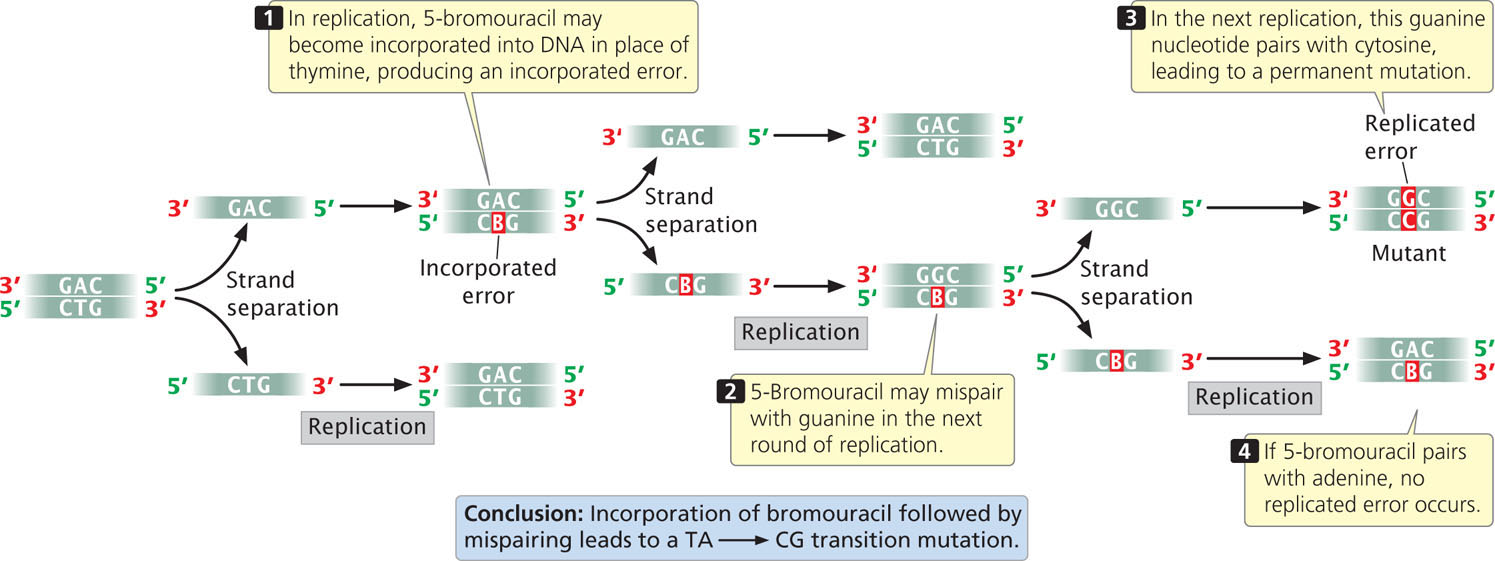
In the laboratory, mutations caused by base analogs can be reversed by treatment with the same analog or by treatment with a different analog.
507
Alkylating Agents
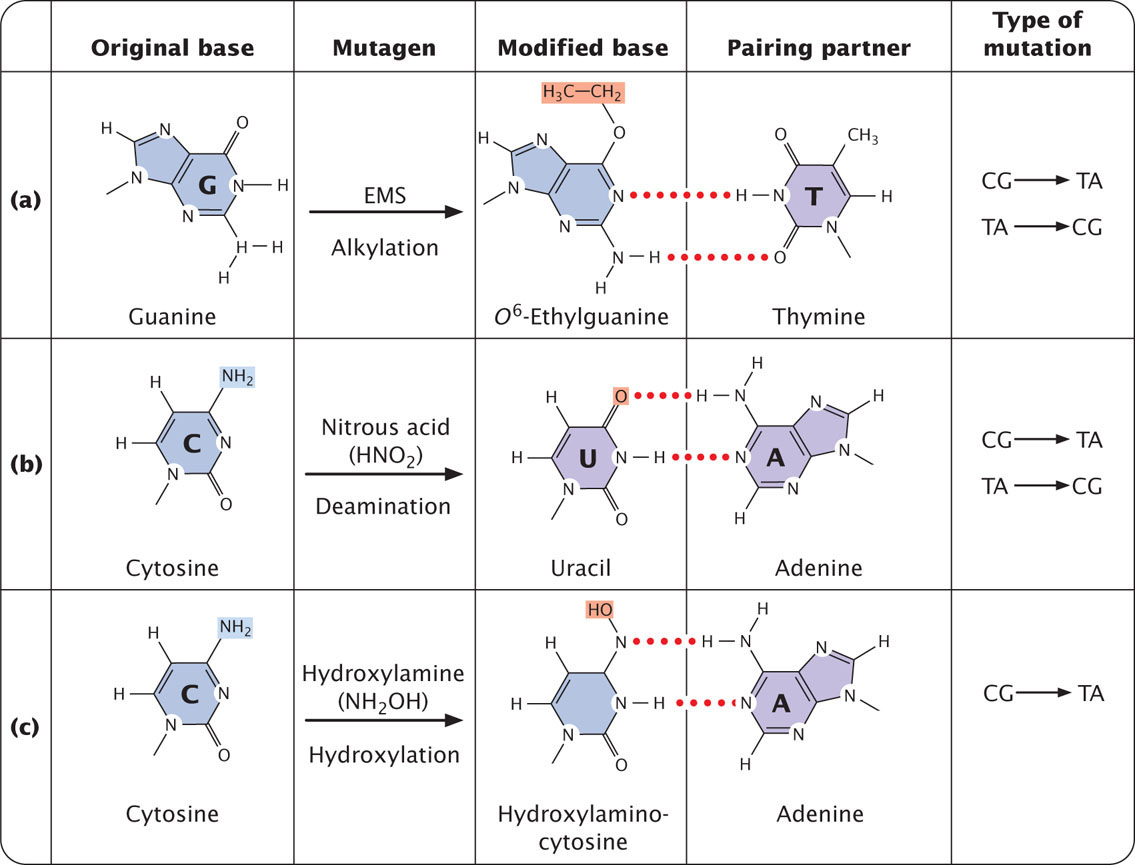
Alkylating agents are chemicals that donate alkyl groups, such as methyl (CH3) and ethyl (CH3–CH2) groups, to nucleotide bases. For example, ethylmethylsulfonate (EMS) adds an ethyl group to guanine, producing O6-ethylguanine, which pairs with thymine (Figure 18.18a). Thus, EMS produces C · G → T · A transitions. EMS is also capable of adding an ethyl group to thymine, producing 4-ethylthymine, which then pairs with guanine, leading to a T · A → C · G transition. Because EMS produces both C · G → T · A and T · A → C · G transitions, mutations produced by EMS can be reversed by additional treatment with EMS. Mustard gas is another alkylating agent.
Deamination
In addition to its spontaneous occurrence (see Figure 18.15), deamination can be induced by some chemicals. For instance, nitrous acid deaminates cytosine, creating uracil, which in the next round of replication pairs with adenine (Figure 18.18b), producing a C · G → T · A transition mutation. Nitrous acid changes adenine into hypoxanthine, which pairs with cytosine, leading to a T · A → C · G transition. Nitrous acid also deaminates guanine, producing xanthine, which pairs with cytosine just as guanine does; however, xanthine can also pair with thymine, leading to a C · G → T · A transition. Nitrous acid produces exclusively transition mutations and, because both C · G → T · A and T · A → C · G transitions are produced, these mutations can be reversed with nitrous acid.
508
Hydroxylamine
Hydroxylamine is a very specific base-modifying mutagen that adds a hydroxyl group to cytosine, converting it into hydroxylaminocytosine (Figure 18.18c). This conversion increases the frequency of a rare tautomer that pairs with adenine instead of guanine and leads to C · G → T · A transitions. Because hydroxylamine acts only on cytosine, it will not generate T · A → C · G transitions; thus, hydroxylamine will not reverse the mutations that it produces. ![]() TRY PROBLEM 25
TRY PROBLEM 25
Oxidative Reactions
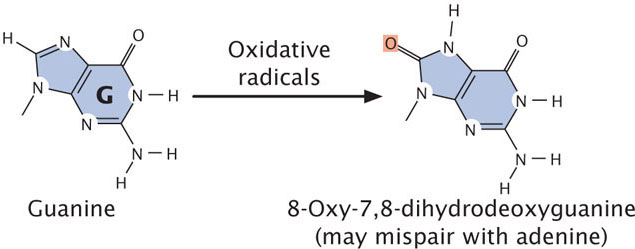
Reactive forms of oxygen (including superoxide radicals, hydrogen peroxide, and hydroxyl radicals) are produced in the course of normal aerobic metabolism, as well as by radiation, ozone, peroxides, and certain drugs. These reactive forms of oxygen damage DNA and induce mutations by bringing about chemical changes in DNA. For example, oxidation converts guanine into 8-oxy-7,8-dihydrodeoxyguanine (Figure 18.19), which frequently mispairs with adenine instead of cytosine, causing a G · C → T · A transversion mutation.
Intercalating Agents
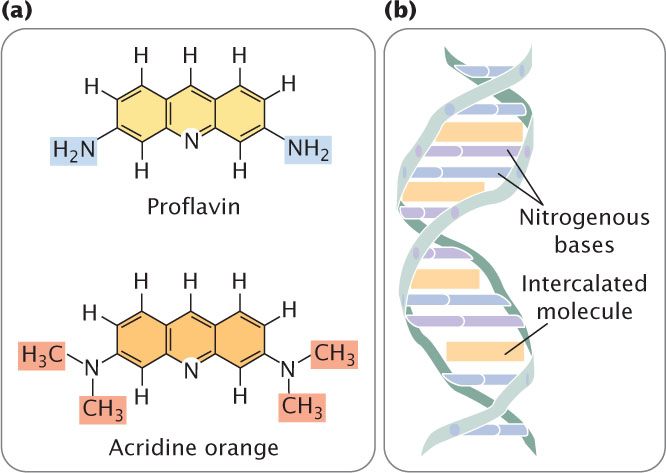
Proflavin, acridine orange, ethidium bromide, and dioxin are intercalating agents (Figure 18.20a), which produce mutations by sandwiching themselves (intercalating) between adjacent bases in DNA, distorting the three-dimensional structure of the helix and causing single-nucleotide insertions and deletions in replication (Figure 18.20b). These insertions and deletions frequently produce frameshift mutations, and so the mutagenic effects of intercalating agents are often severe. Because intercalating agents generate both additions and deletions, they can reverse the effects of their own mutations.
CONCEPTS
Chemicals can produce mutations by a number of mechanisms. Base analogs are incorporated into DNA and frequently pair with the wrong base. Alkylating agents, deaminating chemicals, hydroxylamine, and oxidative radicals change the structure of DNA bases, thereby altering their pairing properties. Intercalating agents wedge between the bases and cause single-base insertions and deletions in replication.
 CONCEPT CHECK 4
CONCEPT CHECK 4
Base analogs are mutagenic because of which characteristic?
- They produce changes in DNA polymerase that cause it to malfunction.
- They distort the structure of DNA.
- They are similar in structure to the normal bases.
- They chemically modify the normal bases.
Radiation
In 1927, Hermann Muller demonstrated that mutations in fruit flies could be induced by X-rays. The results of subsequent studies showed that X-rays greatly increase mutation rates in all organisms. Because of their high energies, X-rays, gamma rays, and cosmic rays are all capable of penetrating tissues and damaging DNA. These forms of radiation, called ionizing radiation, dislodge electrons from the atoms that they encounter, changing stable molecules into free radicals and reactive ions, which then alter the structures of bases and break phosphodiester bonds in DNA. Ionizing radiation also frequently results in double-strand breaks in DNA. Attempts to repair these breaks can produce chromosome mutations (discussed in Chapter 8).
Ultraviolet (UV) light has less energy than does ionizing radiation and does not eject electrons but is nevertheless highly mutagenic. Purine and pyrimidine bases readily absorb UV light, resulting in the formation of chemical bonds between adjacent pyrimidine molecules on the same strand of DNA and in the creation of pyrimidine dimers (Figure 18.21a). Pyrimidine dimers consisting of two thymine bases (called thymine dimers) are most frequent, but cytosine dimers and thymine-cytosine dimers also can form. Dimers distort the configuration of DNA (Figure 18.21b) and often block replication. Most pyrimidine dimers are immediately repaired by mechanisms discussed later in this chapter, but some escape repair and inhibit replication and transcription.
509
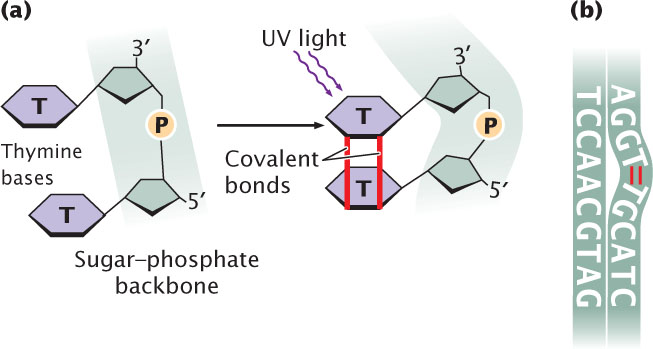
When pyrimidine dimers block replication, cell division is inhibited and the cell usually dies; for this reason, UV light kills bacteria and is an effective sterilizing agent. For a mutation—a hereditary error in the genetic instructions—to occur, the replication block must be overcome. Bacteria can sometimes circumvent replication blocks produced by pyrimidine dimers and other types of DNA damage by means of the SOS system. This system allows replication blocks to be overcome but, in the process, makes numerous mistakes and greatly increases the rate of mutation. Indeed, the very reason that replication can proceed in the presence of a block is that the enzymes in the SOS system do not strictly adhere to the base-pairing rules. The trade-off is that replication can continue and the cell survives, but only by sacrificing the normal accuracy of DNA synthesis.
CONCEPTS
Ionizing radiation such as X-rays and gamma rays damages DNA by dislodging electrons from atoms; these electrons then break phosphodiester bonds and alter the structure of bases. Ultraviolet light causes mutations primarily by producing pyrimidine dimers that disrupt replication and transcription. The SOS system enables bacteria to overcome replication blocks but introduces mistakes in replication.