22.1 Development Takes Place Through Cell Determination
Every multicellular organism begins life as a unicellular, fertilized egg. This single-celled zygote undergoes repeated cell divisions, eventually producing millions or trillions of cells that constitute a complete adult organism. Initially, each cell in the embryo is totipotent: it has the potential to develop into any cell type. Many cells in plants and fungi remain totipotent, but animal cells usually become committed to developing into specific types of cells after just a few early embryonic divisions. This commitment often comes well before a cell begins to exhibit any characteristics of a particular cell type; when the cell becomes committed, it does not normally reverse its fate and develop into a different cell type. A cell becomes committed by a process called determination.
For many years, the work of developmental biologists was limited to describing the changes that take place in the course of development, because techniques for probing the intracellular processes behind these changes were unavailable. But, in recent years powerful genetic and molecular techniques have had a tremendous influence on the study of development; for example, DNA sequencing has provided much information about the nature and organization of DNA sequences that control developmental processes. In some model systems such as Drosophila and Arabidopsis, the molecular mechanisms underlying developmental change are now beginning to be understood.
If all cells in a multicellular organism are derived from the same original cell, how do different cell types arise? Before the 1950s, two hypotheses were considered. One possibility was that throughout development genes might be selectively lost or altered, causing different cell types to have different genomes. Alternatively, each cell might contain the same genetic information, but different genes might be expressed in each cell type. The results of early cloning experiments helped settle this issue.
Cloning Experiments on Plants
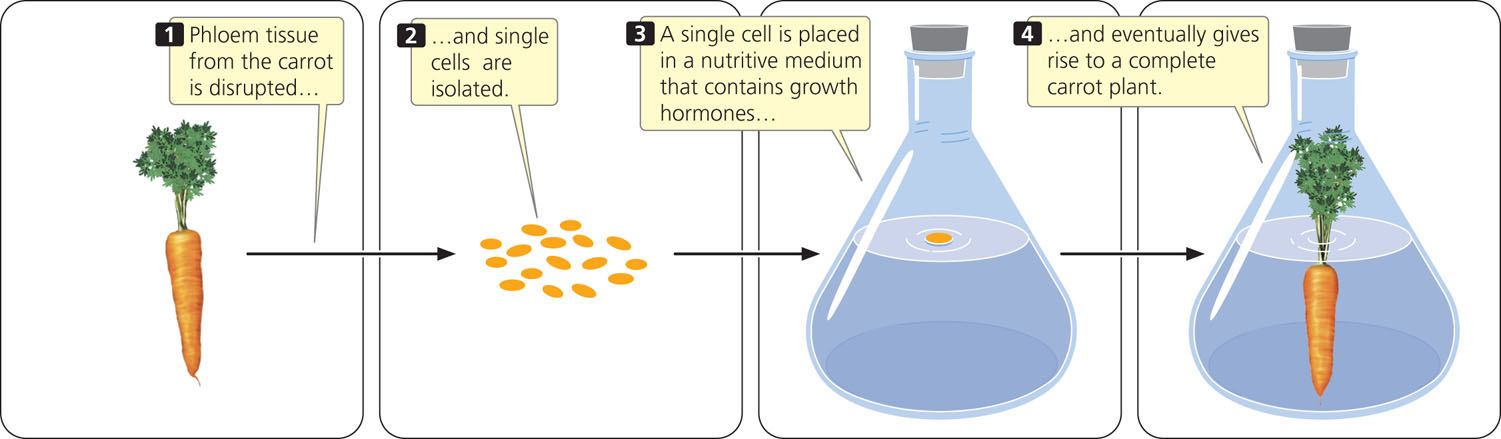
In the 1950s, Frederick Steward developed methods for cloning plants. He disrupted phloem tissue from the root of a carrot by separating and isolating individual cells, and then placed individual cells in a sterile medium that contained nutrients and other substances required for growth. Steward was successful in getting the cells to grow and divide, and eventually he obtained whole edible carrots from single cells (Figure 22.2). Because all parts of the plant were regenerated from a specialized phloem cell, he concluded that each phloem cell contained the genetic potential for a whole plant; none of the original genetic material was lost during determination.
Cloning Experiments on Animals
The results of other studies demonstrated that most animal cells also retain a complete set of genetic information during development. In 1952, Robert Briggs and Thomas King removed the nuclei from unfertilized oocytes of the frog Rana pipiens. They then isolated nuclei from frog blastulas (an early embryonic stage), injected these nuclei individually into the oocytes, and pricked the eggs with a needle to stimulate them to divide. Although most were damaged in the process, a few eggs developed into complete tadpoles that eventually metamorphosed into frogs.
In the late 1960s, John Gurdon used these methods to successfully clone a few frogs with nuclei isolated from the intestinal cells of tadpoles. This suggested that the differentiated intestinal cells carried the genetic information necessary to encode traits found in all other cells. However, Gurdon’s successful clonings may have resulted from the presence of a few undifferentiated stem cells in the intestinal tissue, which were inadvertently used as the nuclei donors.
In 1997, researchers at the Roslin Institute of Scotland announced that they had successfully cloned a sheep by using the genetic material from a differentiated cell of an adult animal. To perform this experiment, they fused an udder cell from a white-faced Finn Dorset ewe with an enucleated egg cell and stimulated the egg electrically to initiate development. After growing the embryo in the laboratory for a week, they implanted it into a Scottish black-faced surrogate mother. Dolly, the first mammal cloned from an adult cell, was born on July 5, 1996 (Figure 22.3). Since then, a number of other animals including sheep, goats, mice, rabbits, cows, pigs, horses, mules, dogs, and cats have been cloned from differentiated adult cells. Importantly, although Dolly and other mammals that have been cloned contain the same nuclear genetic material as that of their cloned parent, they are not identical for cytoplasmic genes, such as those on the mitochondrial chromosome, because the cytoplasm is donated by both the donor cell and the enucleated egg cell.

The cloning experiments demonstrated that genetic material is not lost or permanently altered during development: development must require the selective expression of genes. But how do cells regulate their gene expression in a coordinated manner to give rise to a complex, multicellular organism? Research has now begun to provide some answers to this important question.
CONCEPTS
The ability to clone plants and animals from single specialized cells demonstrates that genes are not lost or permanently altered during development.
 CONCEPT CHECK 1
CONCEPT CHECK 1Scientists have cloned some animals by injecting a nucleus from an early embryo into an enucleated egg cell. Does this outcome demonstrate that genetic material is not lost during development? Why or why not?