25.3 Nonrandom Mating Affects the Genotypic Frequencies of a Population
An assumption of the Hardy–Weinberg law is that mating is random with respect to genotype. Nonrandom mating affects the way in which alleles combine to form genotypes and alters the genotypic frequencies of a population.
We can distinguish between two types of nonrandom mating. Positive assortative mating refers to a tendency for like individuals to mate. For example, humans exhibit positive assortative mating for height: tall people mate preferentially with other tall people; short people mate preferentially with other short people. Negative assortative mating refers to a tendency for unlike individuals to mate. If people engaged in negative assortative mating for height, tall and short people would preferentially mate. Assortative mating is usually for a particular trait and will affect only those genes that encode the trait (and genes closely linked to them).
One form of nonrandom mating is inbreeding, which is preferential mating between related individuals. Inbreeding is actually positive assortative mating for relatedness, but it differs from other types of assortative mating because it affects all genes, not just those that determine the trait for which the mating preference exists. Inbreeding causes a departure from the Hardy–Weinberg equilibrium frequencies of p2, 2pq, and q2. More specifically, it leads to an increase in the proportion of homozygotes and a decrease in the proportion of heterozygotes in a population. Outcrossing is the avoidance of mating between related individuals.
In a diploid organism, a homozygous individual has two copies of the same allele. These two copies may be the same in state, which means that the two alleles are alike in structure and function but do not have a common origin. Alternatively, the two alleles in a homozygous individual may be the same because they are identical by descent; that is, the copies are descended from a single allele that was present in an ancestor (Figure 25.4). If we go back far enough in time, many alleles are likely to be identical by descent, but for calculating the effects of inbreeding we consider identity by descent by going back only a few generations.
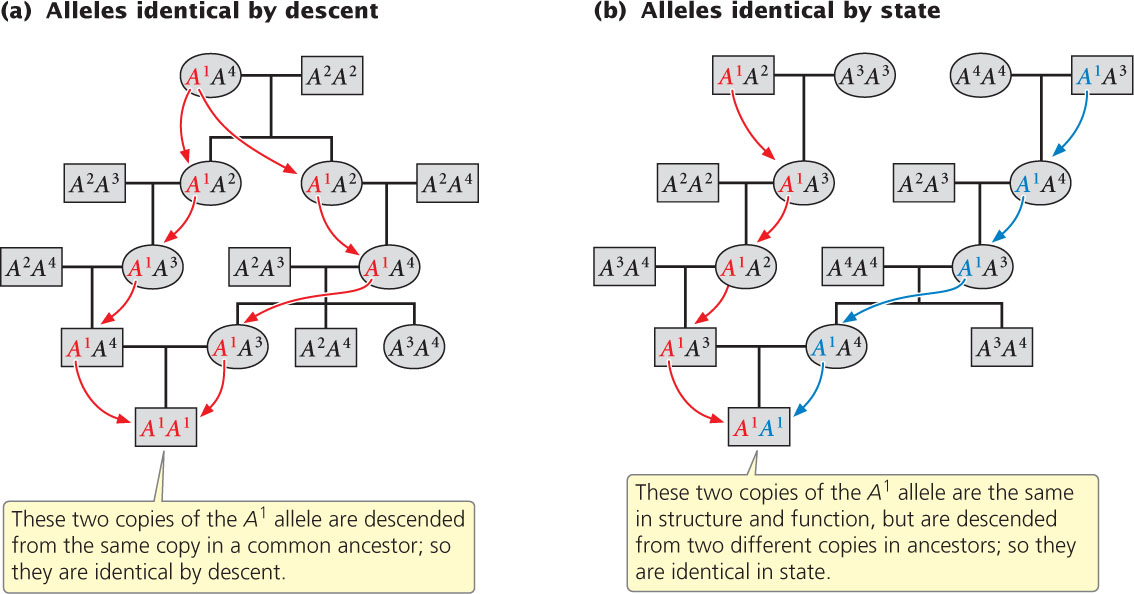
Inbreeding is usually measured by the inbreeding coefficient, designated F, which is a measure of the probability that two alleles are identical by descent. Inbreeding coefficients can range from 0 to 1. A value of 0 indicates that mating in a large population is random; a value of 1 indicates that all alleles are identical by descent. Inbreeding coefficients can be calculated from analyses of pedigrees or they can be determined from the reduction in the heterozygosity of a population. Although we will not go into the details of how F is calculated, an understanding of how inbreeding affects genotypic frequencies is important.
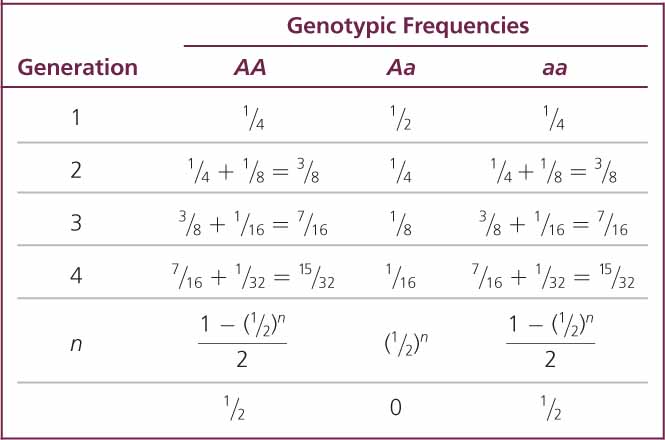
When inbreeding takes place, the proportion of heterozygotes decreases by 2Fpq, and half of this value (Fpq) is added to the proportion of each homozygote each generation. The frequencies of the genotypes will then be
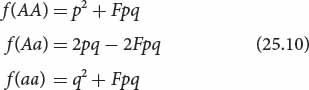
Consider a population that reproduces by self-fertilization (F = 1). We will assume that this population begins with genotypic frequencies in Hardy–Weinberg proportions (p2, 2pq, and q2). With selfing, each homozygote produces progeny only of the same homozygous genotype (AA × AA produces all AA; and aa × aa produces all aa), whereas only half the progeny of a heterozygote will be like the parent (Aa × Aa produces 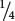 AA,
AA, 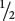 Aa, and
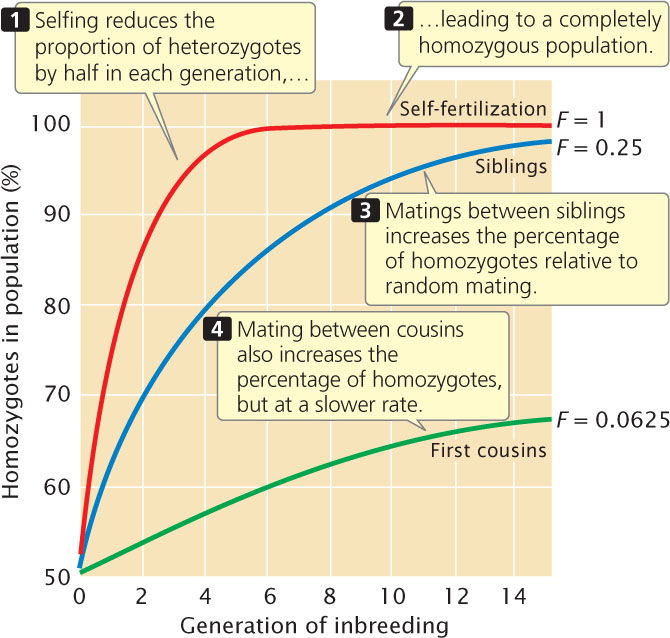
aa). Selfing therefore reduces the proportion of heterozygotes in the population by half with each generation, until all genotypes in the population are homozygous (Table 25.2 and Figure 25.5). Although all individuals in the population are homozygous at this point, both types of homozygotes (AA and aa) are still present, provided neither homozygote has an advantage over the other (there are no selective differences).
Aa, and
aa). Selfing therefore reduces the proportion of heterozygotes in the population by half with each generation, until all genotypes in the population are homozygous (Table 25.2 and Figure 25.5). Although all individuals in the population are homozygous at this point, both types of homozygotes (AA and aa) are still present, provided neither homozygote has an advantage over the other (there are no selective differences). ![]() TRY PROBLEM 28
TRY PROBLEM 28
724
For most outcrossing species, close inbreeding is harmful because it increases the proportion of homozygotes and thereby boosts the probability that deleterious and lethal recessive alleles will combine to produce homozygotes with a harmful trait. Assume that a recessive allele (a) that causes a genetic disease has a frequency (q) of 0.01. If the population mates randomly (F = 0), the frequency of individuals affected with the disease (aa) will be q2 = 0.012 = 0.0001; so only 1 in 10,000 individuals will have the disease. However, if F = 0.25 (the equivalent of brother–sister mating), then the expected frequency of the homozygote genotype is q2 + Fpq = (0.01)2 + (0.25)(0.99)(0.01) = 0.0026; thus, the genetic disease is 26 times as frequent at this level of inbreeding. This increased appearance of lethal and deleterious traits with inbreeding is termed inbreeding depression; the more intense the inbreeding, the more severe the inbreeding depression. To see the effects of inbreeding on genotypic frequencies, view the Mini-Tutorial in  Animation 25.1.
Animation 25.1.
725
The harmful effects of inbreeding have been recognized for thousands of years and may be the basis of cultural taboos against mating between close relatives. William Schull and James Neel found that for each 10% increase in F the mean IQ of Japanese children dropped six points. Child mortality also increases with close inbreeding (Table 25.3); children of first cousins have a 40% increase in mortality over that seen among the children of unrelated people. Inbreeding also has deleterious effects on crops (Figure 25.6) and domestic animals.
| Genetic Relationship of Parents | F | Mortality of Children (through 12 years of age) |
|---|---|---|
| Unrelated | 0 | 0.082 |
| Second cousins | 0.016 (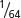 ) ) |
0.108 |
| First cousins | 0.0625 (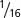 ) ) |
0.114 |
| Source: After D. L. Hartl and A. G. Clark, Principles of Population Genetics, 2d ed. (Sunderland, Mass.: Sinauer, 1989), Table 2. Original data from W. J. Schull and J. V. Neel, The Effects of Inbreeding on Japanese Children (New York: Harper & Row, 1965). | ||
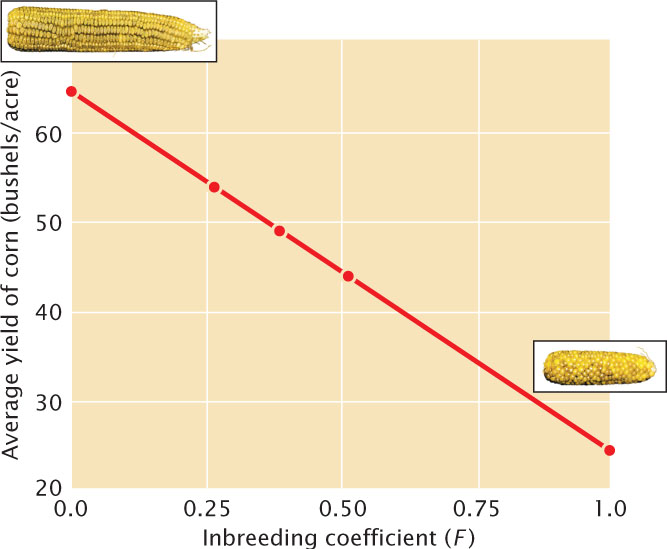
Inbreeding depression is most often studied in humans, as well as in plants and animals reared in captivity, but the negative effects of inbreeding may be more severe in natural populations. Julie Jimenez and her colleagues collected wild mice from a natural population in Illinois and bred them in the laboratory for three to four generations. Laboratory matings were chosen so that some mice had no inbreeding, whereas others had an inbreeding coefficient of 0.25. When both types of mice were released back into the wild, the weekly survival of the inbred mice was only 56% of that of the noninbred mice. Inbred male mice also continously lost body weight after release into the wild, whereas noninbred male mice initially lost weight but then regained it within a few days after release.
In spite of the fact that inbreeding is generally harmful for outcrossing species, a number of plants and animals regularly inbreed and are successful (Figure 25.7). As stated earlier, inbreeding increases homozygosity, and eventually all individuals in the population become homozygous. If a species undergoes inbreeding for a number of generations, many deleterious recessive alleles are weeded out by natural or artificial selection so that the population becomes homozygous for beneficial alleles. In this way, the harmful effects of inbreeding may eventually be eliminated, leaving a population that is homozygous for beneficial traits.

CONCEPTS
Nonrandom mating alters the frequencies of the genotypes but not the frequencies of the alleles. Inbreeding is preferential mating between related individuals. With inbreeding, the frequency of homozygotes increases, whereas the frequency of heterozygotes decreases.
 CONCEPT CHECK 5
CONCEPT CHECK 5
What is the effect of outcrossing on a population?
- Allelic frequencies change.
- There will be more heterozygotes than predicted by the Hardy–Weinberg law.
- There will be fewer heterozygotes than predicted by the Hardy–Weinberg law.
- Genotypic frequencies will equal those predicted by the Hardy–Weinberg law.
726