5.1 Additional Factors at a Single Locus Can Affect the Results of Genetic Crosses
In Chapter 3, we learned that the principle of segregation and the principle of independent assortment enable us to predict the outcomes of genetic crosses. Here, we examine several additional factors acting at individual loci that can alter the phenotypic ratios predicted by Mendel’s principles.
Types of Dominance
One of Mendel’s important contributions to the study of heredity is the concept of dominance—the idea that an individual organism possesses two different alleles for a characteristic but the trait encoded by only one of the alleles is observed in the phenotype. With dominance, the heterozygote possesses the same phenotype as one of the homozygotes.
Mendel observed dominance in all of the traits that he chose to study extensively, but he was aware that not all characteristics exhibit dominance. He conducted some crosses concerning the length of time that pea plants take to flower. For example, when he crossed two homozygous varieties that differed in their flowering time by an average of 20 days, the length of time taken by the F1 plants to flower was intermediate between those of the two parents. When the heterozygote has a phenotype intermediate between the phenotypes of the two homozygotes, the trait is said to display incomplete dominance.
Complete and Incomplete DominanceDominance can be understood in regard to how the phenotype of the heterozygote relates to the phenotypes of the homozygotes. In the example presented in the upper panel of Figure 5.1, flower color potentially ranges from red to white. One homozygous genotype, A1A1, produces red pigment, resulting in red flowers; another, A2A2, produces no pigment, resulting in white flowers. Where the heterozygote falls in the range of phenotypes determines the type of dominance. If the heterozygote (A1 A2) produces the same amount of pigment as the A1A1 homozygote, resulting in red, then the A1 allele displays complete dominance over the A2 allele; that is, red is dominant over white. If, on the other hand, the heterozygote produces no pigment, resulting in flowers with the same color as the A2A2 homozygote (white), then the A2 allele is completely dominant, and white is dominant over red.
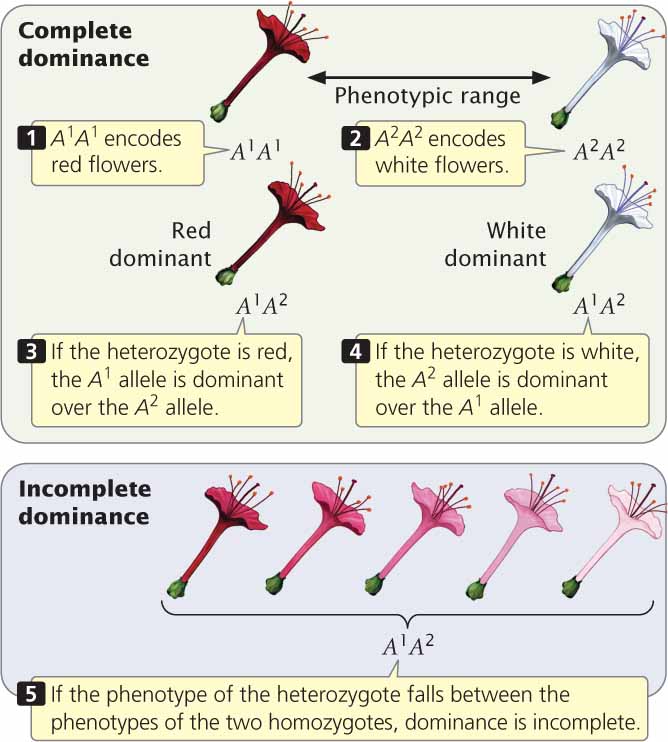
When the heterozygote falls in between the phenotypes of the two homozygotes, dominance is incomplete. With incomplete dominance, the heterozygote need not be exactly intermediate between the two homozygotes (see the bottom panel of Figure 5.1); it might be a slightly lighter shade of red or a slightly pink shade of white. As long as the heterozygote’s phenotype can be differentiated and falls within the range of the two homozygotes, dominance is incomplete.
Incomplete dominance is also exhibited in the fruit color of eggplant. When a homozygous plant that produces purple fruit (PP) is crossed with a homozygous plant that produces white fruit (pp), all the heterozygous F1 (Pp) produce violet fruit (Figure 5.2a). When the F1 are crossed with each other, 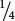 of the F2 are purple (PP),
of the F2 are purple (PP), 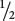 are violet (Pp), and
are white (pp), as shown in Figure 5.2b. Note that this 1 : 2 : 1 ratio is different from the 3 : 1 ratio that we would observe if eggplant fruit color exhibited complete dominance. Another example of incomplete dominance is feather color in chickens. A cross between a homozygous black chicken and a homozygous white chicken produces F1 chickens that are gray. If these gray F1 are intercrossed, they produce F2 birds in a ratio of 1 black : 2 gray : 1 white.
are violet (Pp), and
are white (pp), as shown in Figure 5.2b. Note that this 1 : 2 : 1 ratio is different from the 3 : 1 ratio that we would observe if eggplant fruit color exhibited complete dominance. Another example of incomplete dominance is feather color in chickens. A cross between a homozygous black chicken and a homozygous white chicken produces F1 chickens that are gray. If these gray F1 are intercrossed, they produce F2 birds in a ratio of 1 black : 2 gray : 1 white.
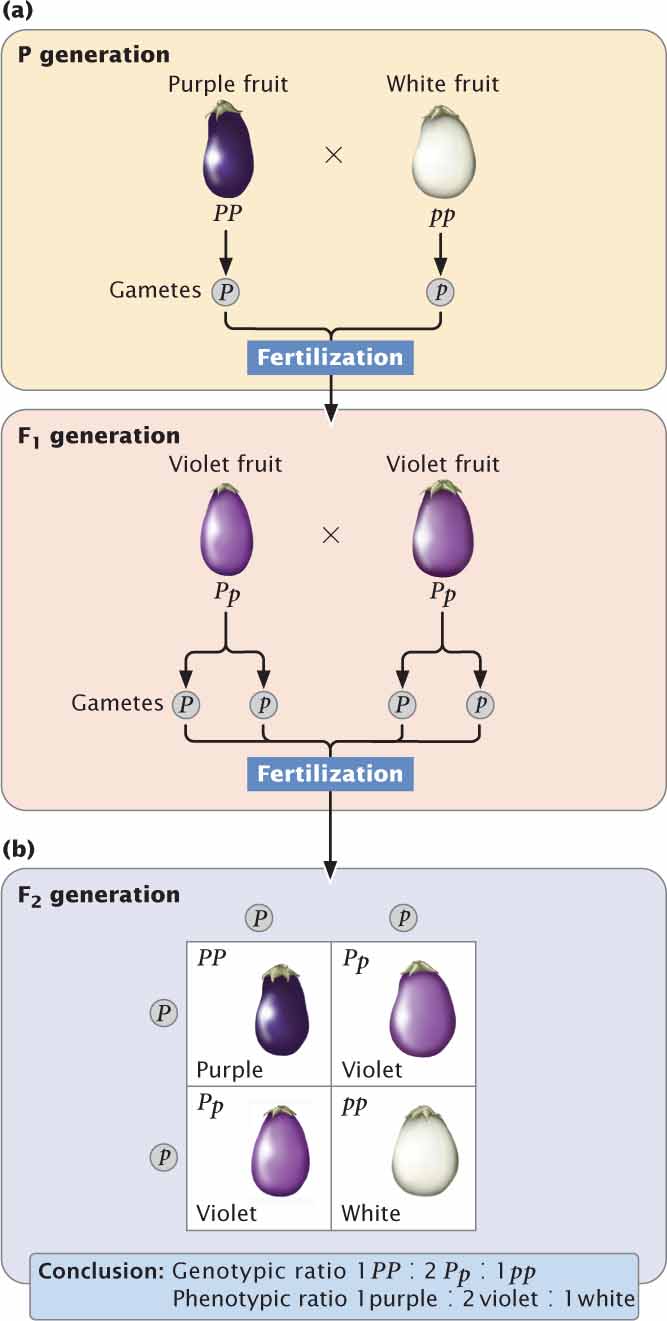
We should now add the 1 : 2 : 1 ratio to those phenotypic ratios for simple crosses presented in Chapter 3 (see Table 3.5). A 1 : 2 : 1 phenotypic ratio arises in the progeny of a cross between two parents heterozygous for a character that exhibits incomplete dominance (Aa × Aa). The genotypic ratio among these progeny also is 1 : 2 : 1. When a trait displays incomplete dominance, the genotypic ratios and phenotypic ratios of the offspring are the same, because each genotype has its own phenotype. The important thing to remember about dominance is that it affects the way the genes are expressed (the phenotype) but not the way the genes are inherited.
CONCEPTS
Incomplete dominance is exhibited when the heterozygote has a phenotype intermediate between the phenotypes of the two homozygotes. When a trait exhibits incomplete dominance, a cross between two heterozygotes produces a 1 : 2 : 1 phenotypic ratio in the progeny.
 CONCEPT CHECK 1
CONCEPT CHECK 1If an F1 eggplant in Figure 5.2 is used in a testcross, what proportion of the progeny from this cross will be white?
- All the progenyc.
-
-
- 0
CodominanceAnother type of interaction between alleles is codominance, in which the phenotype of the heterozygote is not intermediate between the phenotypes of the homozygotes; rather, the heterozygote simultaneously expresses the phenotypes of both homozygotes. An example of codominance is seen in the MN blood types.
The MN locus encodes one of the types of antigens on red blood cells. Unlike antigens foreign to the ABO and Rh blood groups (which also encode red-blood-cell antigens), foreign MN antigens do not elicit a strong immunological reaction; therefore, the MN blood types are not routinely considered in blood transfusions. At the MN locus, there are two alleles: the LM allele, which encodes the M antigen; and the LN allele, which encodes the N antigen. Homozygotes with genotype LMLM express the M antigen on their red blood cells and have the M blood type. Homozygotes with genotype LNLN express the N antigen and have the N blood type. Heterozygotes with genotype LMLN exhibit codominance and express both the M and the N antigens; they have blood-type MN.
Some students might ask why the pink flowers illustrated in the bottom panel of Figure 5.1 exhibit incomplete dominance—that is, why is this outcome not an example of codominance? The flowers would exhibit codominance only if the heterozygote produced both red and white pigments, which then combined to produce a pink phenotype. However, in our example, the heterozygote produces only red pigment. The pink phenotype comes about because the amount of pigment produced by the heterozygote is less than the amount produced by the A1A1 homozyogote. So, here, the alleles clearly exhibit incomplete dominance, not codominance. The differences between complete dominance, incomplete dominance, and codominance are summarized in Table 5.1. ![]() TRY PROBLEM 13
TRY PROBLEM 13
| Type of Dominance | Definition |
|---|---|
| Complete dominance | phenotype of the heterozygote is the same as the phenotype of one of the homozygotes. |
| Incomplete dominance | phenotype of the heterozygote is intermediate (falls within the range) between the phenotypes of the two homozygotes. |
| Codominance | phenotype of the heterozygote includes the phenotypes of both homozygotes. |
Level Of Phenotype Observed May Affect DominancePhenotypes can frequently be observed at several different levels, including the anatomical level, the physiological level, and the molecular level. The type of dominance exhibited by a characteristic depends on the level of the phenotype examined. This dependency is seen in cystic fibrosis, a common genetic disorder in Caucasians and usually considered to be a recessive disease. People who have cystic fibrosis produce large quantities of thick, sticky mucus, which plugs up the airways of the lungs and clogs the ducts leading from the pancreas to the intestine, causing frequent respiratory infections and digestive problems. Even with medical treatment, patients with cystic fibrosis suffer chronic, life-threatening medical problems.
The gene responsible for cystic fibrosis resides on the long arm of chromosome 7. It encodes a protein termed cystic fibrosis transmembrane conductance regulator (CFTR), which acts as a gate in the cell membrane and regulates the movement of chloride ions into and out of the cell. Persons with cystic fibrosis have a mutated, dysfunctional form of CFTR that causes the channel to stay closed, and so chloride ions build up in the cell. This buildup causes the formation of thick mucus and produces the symptoms of the disease.
Most people have two copies of the normal allele for CFTR and produce only functional CFTR protein. Those with cystic fibrosis possess two copies of the mutated CFTR allele and produce only the defective CFTR protein. Heterozygotes, having one normal and one defective CFTR allele, produce both functional and defective CFTR protein. Thus, at the molecular level, the alleles for normal and defective CFTR are codominant because both alleles are expressed in the heterozygote. However, because one functional allele produces enough functional CFTR protein to allow normal chloride-ion transport, the heterozygote exhibits no adverse effects, and the mutated CFTR allele appears to be recessive at the physiological level. The type of dominance expressed by an allele, as illustrated in this example, is a function of the phenotypic aspect of the allele that is observed.
Characteristics Of DominanceSeveral important characteristics of dominance should be emphasized. First, dominance is a result of interactions between genes at the same locus; in other words, dominance is allelic interaction. Second, dominance does not alter the way in which the genes are inherited; it only influences the way in which they are expressed as a phenotype. The allelic interaction that characterizes dominance is therefore interaction between the products of the genes. Finally, dominance is frequently “in the eye of the beholder,” meaning that the classification of dominance depends on the level at which the phenotype is examined. As seen for cystic fibrosis, an allele may exhibit codominance at one level and be recessive at another level.
CONCEPTS
Dominance entails interactions between genes at the same locus (allelic genes) and is an aspect of the phenotype; dominance does not affect the way in which genes are inherited. The type of dominance exhibited by a characteristic frequently depends on the level of the phenotype examined.
CONCEPT CHECK 2How do complete dominance, incomplete dominance, and codominance differ?
Penetrance and Expressivity
In the genetic crosses presented thus far, we have considered only the interactions of alleles and have assumed that every individual organism having a particular genotype expresses the expected phenotype. We assumed, for example, that the genotype Rr always produces round seeds and that the genotype rr always produces wrinkled seeds. For some characters, however, such an assumption is incorrect: the genotype does not always produce the expected phenotype, a phenomenon termed incomplete penetrance.
Incomplete penetrance is seen in human polydactyly, the condition of having extra fingers or toes (Figure 5.3). There are several different forms of human polydactyly, but the trait is usually caused by a dominant allele. Occasionally, people possess the allele for polydactyly (as evidenced by the fact that their children inherit the polydactyly) but nevertheless have a normal number of fingers and toes. In these cases, the gene for polydactyly is not fully penetrant. Penetrance is defined as the percentage of individual organisms having a particular genotype that express the expected phenotype. For example, if we examined 42 people having an allele for polydactyly and found that only 38 of them were polydactylous, the penetrance would be 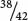 = 0.90 (90%).
= 0.90 (90%).
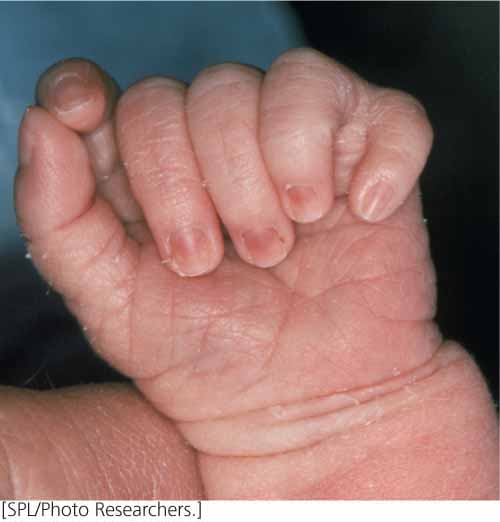
A related concept is that of expressivity, the degree to which a trait is expressed. In addition to incomplete penetrance, polydactyly exhibits variable expressivity. Some polydactylous persons possess extra fingers or toes that are fully functional, whereas others possess only a small tag of extra skin.
Incomplete penetrance and variable expressivity are due to the effects of other genes and to environmental factors that can alter or completely suppress the effect of a particular gene. For example, a gene may encode an enzyme that produces a particular phenotype only within a limited temperature range. At higher or lower temperatures, the enzyme does not function and the phenotype is not expressed; the allele encoding such an enzyme is therefore penetrant only within a particular temperature range (also see Environmental Effects on the Phenotype). Many characters exhibit incomplete penetrance and variable expressivity; thus the mere presence of a gene does not guarantee its expression. ![]() TRY PROBLEM 15
TRY PROBLEM 15
CONCEPTS
Penetrance is the percentage of individuals having a particular genotype that express the associated phenotype. Expressivity is the degree to which a trait is expressed. Incomplete penetrance and variable expressivity result from the influence of other genes and environmental factors on the phenotype.
CONCEPT CHECK 3How does incomplete dominance differ from incomplete penetrance?
- Incomplete dominance refers to alleles at the same locus; incomplete penetrance refers to alleles at different loci.
- Incomplete dominance ranges from 0% to 50%; incomplete penetrance ranges from 51% to 99%.
- In incomplete dominance, the heterozygote is intermediate between the homozygotes; in incomplete penetrance, heterozygotes express phenotypes of both homozygotes.
- In incomplete dominance, the heterozygote is intermediate between the homozygotes; in incomplete penetrance, some individuals do not express the expected phenotype.
Lethal Alleles
A lethal allele causes death at an early stage of development—often before birth—and so some genotypes may not appear among the progeny. An example of a lethal allele, originally described by Erwin Baur in 1907, is found in snapdragons. The aurea strain in these plants has yellow leaves. When two plants with yellow leaves are crossed, 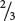 of the progeny have yellow leaves and
of the progeny have yellow leaves and 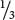 have green leaves. When green is crossed with green, all the progeny have green leaves; however, when yellow is crossed with green,
of the progeny have green leaves and
have yellow leaves, confirming that all yellow-leaved snapdragons are heterozygous.
have green leaves. When green is crossed with green, all the progeny have green leaves; however, when yellow is crossed with green,
of the progeny have green leaves and
have yellow leaves, confirming that all yellow-leaved snapdragons are heterozygous.
Another example of a lethal allele is one that determines yellow coat color in mice. A cross between two yellow heterozygous mice produces an initial genotypic ratio of
YY,
Yy, and
yy, but the homozygous YY mice die early in development and do not appear among the progeny, resulting in a 2 : 1 ratio of Yy (yellow) to yy (nonyellow) in offspring (Figure 5.4). A 2 : 1 ratio is almost always produced by a recessive lethal allele; so observing this ratio among the progeny of a cross between individuals with the same phenotype is a strong clue that one of the alleles is lethal. In this example, like that of yellow leaves in snapdragons, the lethal allele is recessive because it causes death only in homozygotes. Unlike its effect on survival, the effect of the allele on color is dominant; in both mice and snapdragons, a single copy of the allele in the heterozygote produces a yellow color. This example illustrates the point made earlier that the type of dominance depends on the aspect of the phenotype examined.
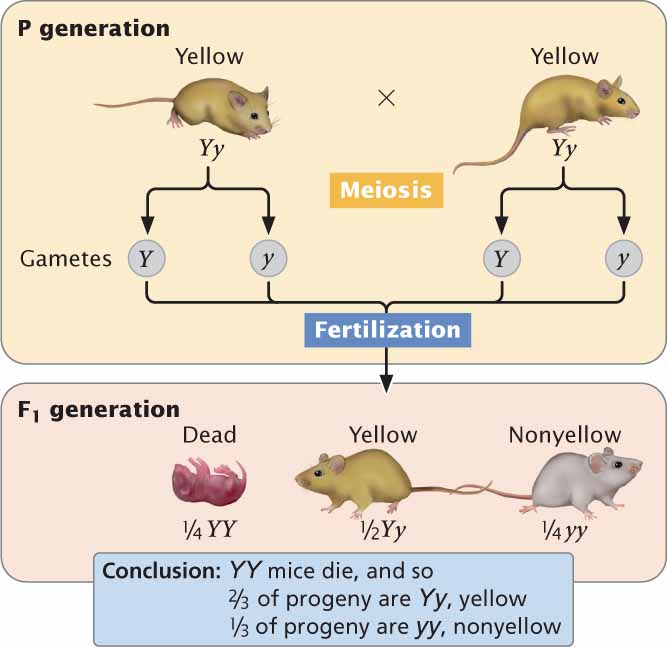
Many lethal alleles in nature are recessive, but lethal alleles can also be dominant; in this case, homozygotes and heterozygotes for the allele die. Truly dominant lethal alleles cannot be transmitted unless they are expressed after the onset of reproduction. ![]() TRY PROBLEM 17
TRY PROBLEM 17
CONCEPTS
A lethal allele causes death, frequently at an early developmental stage, so one or more genotypes are missing from the progeny of a cross. Lethal alleles modify the ratio of progeny resulting from a cross.
CONCEPT CHECK 4A cross between two green corn plants yields
progeny that are green and
progeny that are yellow. What is the genotype of the green progeny?
- WW
- Ww
- ww
- W_ (WW and Ww)
Multiple Alleles
Most of the genetic systems that we have examined so far consist of two alleles. In Mendel’s peas, for instance, one allele encoded round seeds and another encoded wrinkled seeds; in cats, one allele produced a black coat and another produced a gray coat. For some loci, more than two alleles are present within a group of organisms—the locus has multiple alleles (multiple alleles may also be referred to as an allelic series). Although there may be more than two alleles present within a group of organisms, the genotype of each individual diploid organism still consists of only two alleles. The inheritance of characteristics encoded by multiple alleles is no different from the inheritance of characteristics encoded by two alleles, except that a greater variety of genotypes and phenotypes are possible.
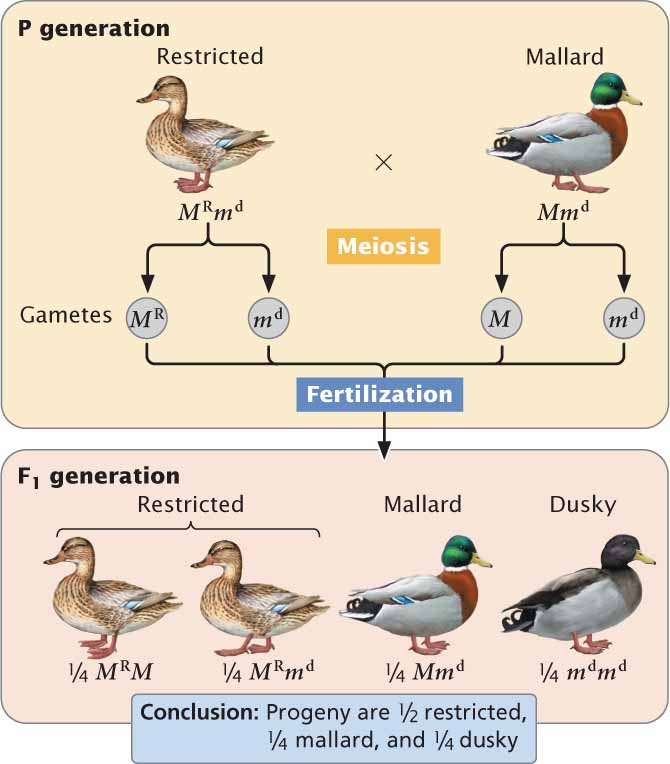
Duck-Feather PatternsAn example of multiple alleles is at a locus that determines the feather pattern of mallard ducks. One allele, M, produces the wild-type mallard pattern. A second allele, MR, produces a different pattern called restricted, and a third allele, md, produces a pattern termed dusky. In this allelic series, restricted is dominant over mallard and dusky, and mallard is dominant over dusky: MR > M > md. The six genotypes possible with these three alleles and their resulting phenotypes are:
| Genotype | Phenotype |
|---|---|
| MRMR | restricted |
| MRM | restricted |
| MRmd | restricted |
| MM | mallard |
| Mmd | mallard |
| mdmd | dusky |
In general, the number of genotypes possible will be [n(n + 1)/2] where n equals the number of different alleles at a locus. Working crosses with multiple alleles is no different from working crosses with two alleles; Mendel’s principle of segregation still holds, as shown in the cross between a restricted duck and a mallard duck (Figure 5.5). ![]() TRY PROBLEM 19
TRY PROBLEM 19
The ABO Blood GroupAnother multiple-allele system is at the locus for the ABO blood group. This locus determines your ABO blood type and, like the MN locus, encodes antigens on red blood cells. The three common alleles for the ABO blood group locus are: IA, which encodes the A antigen; IB, which encodes the B antigen; and i, which encodes no antigen (O). We can represent the dominance relations among the ABO alleles as follows: IA > i, IB > i, IA = IB. Both the IA and the IB alleles are dominant over i and are codominant with each other; the AB phenotype is due to the presence of an IA allele and an IB allele, which results in the production of A and B antigens on red blood cells. A person with genotype ii produces neither antigen and has blood type O. The six common genotypes at this locus and their phenotypes are shown in Figure 5.6a.
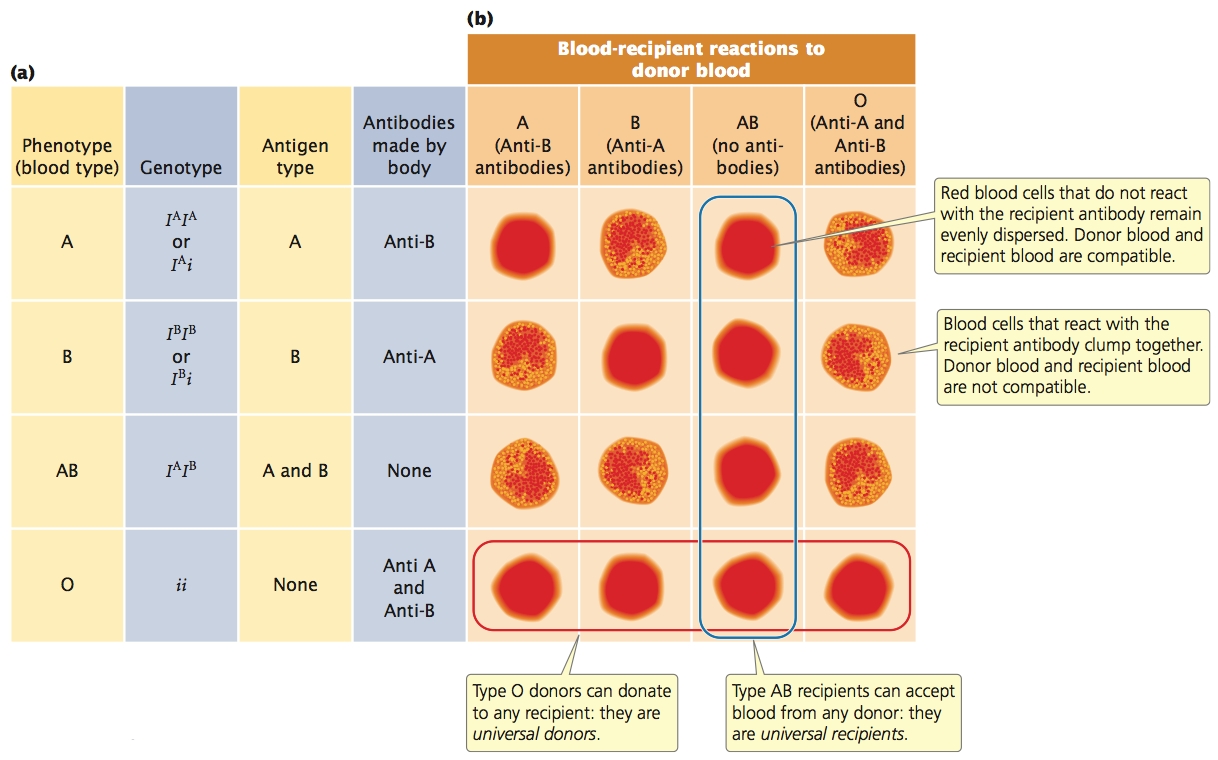
Antibodies are produced against any foreign antigens (see Figure 5.6a). For instance, a person having blood-type A produces anti-B antibodies, because the B antigen is foreign. A person having blood-type B produces anti-A antibodies, and someone having blood-type AB produces neither anti-A nor anti-B antibodies, because neither A nor B antigen is foreign. A person having blood-type O possesses no A or B antigens; consequently, that person produces both anti-A antibodies and anti-B antibodies. The presence of antibodies against foreign ABO antigens means that successful blood transfusions are possible only between persons with certain compatible blood types (Figure 5.6b).
The inheritance of alleles at the ABO locus is illustrated by a paternity suit against the famous movie actor Charlie Chaplin. In 1941, Chaplin met a young actress named Joan Barry, with whom he had an affair. The affair ended in February 1942 but, 20 months later, Barry gave birth to a baby girl and claimed that Chaplin was the father. Barry then sued for child support. At this time, blood typing had just come into widespread use, and Chaplin’s attorneys had Chaplin, Barry, and the child blood typed. Barry had blood-type A, her child had blood-type B, and Chaplin had blood-type O. Could Chaplin have been the father of Barry’s child?
Your answer should be no. Joan Barry had blood-type A, which can be produced by either genotype IAIA or genotype IAi. Her baby possessed blood-type B, which can be produced by either genotype IBIB or genotype IBi. The baby could not have inherited the IB allele from Barry (Barry could not carry an IB allele if she were blood-type A); therefore the baby must have inherited the i allele from her. Barry must have had genotype IAi, and the baby must have had genotype IBi. Because the baby girl inherited her i allele from Barry, she must have inherited the IB allele from her father. Having blood-type O, produced only by genotype ii, Chaplin could not have been the father of Barry’s child. Although blood types can be used to exclude the possibility of paternity (as in this case), they cannot prove that a person is the parent of a child, because many different people have the same blood type.
In the course of the trial to settle the paternity suit against Chaplin, three pathologists testified that it was genetically impossible for Chaplin to have fathered the child. Nevertheless, the jury ruled that Chaplin was the father and ordered him to pay child support and Barry’s legal expenses. ![]() TRY PROBLEM 24
TRY PROBLEM 24
Compound HeterozygotesDifferent alleles often give rise to the same phenotype. For example, cystic fibrosis arises from defects in alleles at the CFTR locus, which encodes a protein that controls the movement of chloride ions into and out of the cell. Over 1000 different alleles at the CFTR locus have been discovered worldwide that can cause cystic fibrosis. Because cystic fibrosis is an autosomal recessive condition, one must normally inherit two defective CFTR alleles to have cystic fibrosis. In some people with cystic fibrosis, these two defective alleles are identical, meaning that the person is homozygous. Others with cystic fibrosis are heterozygous, possessing two different defective alleles. An individual who carries two different alleles at a locus resulting in a recessive phenotype is referred to as compound heterozygote.
CONCEPTS
More than two alleles (multiple alleles) may be present within a group of individual organisms, although each individual diploid organism still has only two alleles at that locus. A compound heterozygote possesses two different alleles that result in a recessive phenotype.
CONCEPT CHECK 5How many genotypes are possible at a locus with five alleles?
- 30
- 27
- 15
- 5