8.3 Aneuploidy Is an Increase or Decrease in the Number of Individual Chromosomes
In addition to chromosome rearrangements, chromosome mutations include changes in the number of chromosomes. Variations in chromosome number can be classified into two basic types: aneuploidy, which is a change in the number of individual chromosomes, and polyploidy, which is a change in the number of chromosome sets.
Aneuploidy can arise in several ways. First, a chromosome may be lost in the course of mitosis or meiosis if, for example, its centromere is deleted. Loss of the centromere prevents the spindle fibers from attaching; so the chromosome fails to move to the spindle pole and does not become incorporated into a nucleus after cell division. Second, the small chromosome generated by a Robertsonian translocation may be lost in mitosis or meiosis. Third, aneuploids may arise through nondisjunction, the failure of homologous chromosomes or sister chromatids to separate in meiosis or mitosis. Nondisjunction leads to some gametes or cells that contain an extra chromosome and other gametes or cells that are missing a chromosome (Figure 8.19). ![]() TRY PROBLEM 29
TRY PROBLEM 29
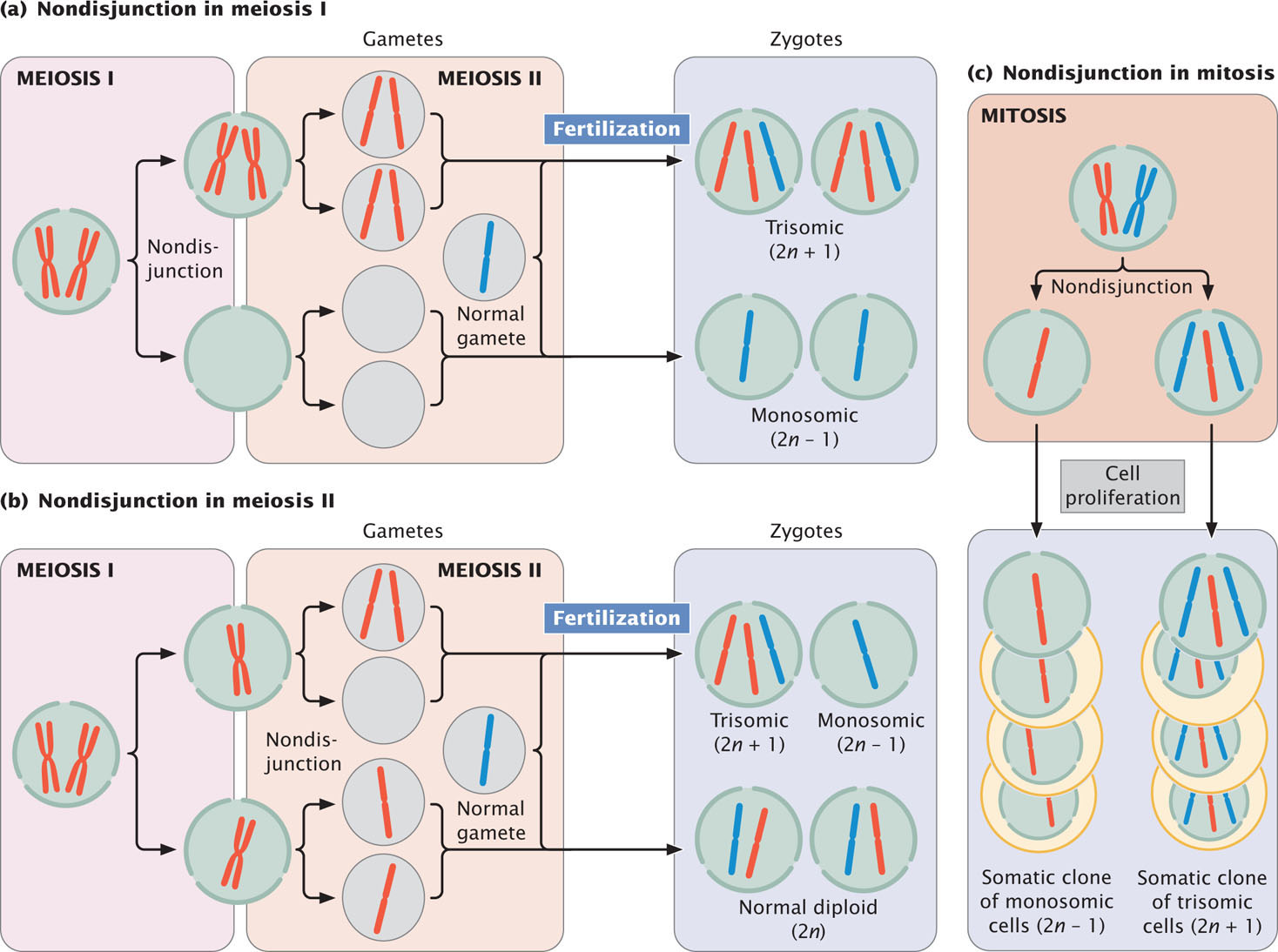
Types of Aneuploidy
We will consider four types of common aneuploid conditions in diploid individuals: nullisomy, monosomy, trisomy, and tetrasomy.
- 1. Nullisomy is the loss of both members of a homologous pair of chromosomes. It is represented as 2n − 2, where n refers to the haploid number of chromosomes. Thus, among humans, who normally possess 2n = 46 chromosomes, a nullisomic zygote has 44 chromosomes.
- 2. Monosomy is the loss of a single chromosome, represented as 2n − 1. A human monosomic zygote has 45 chromosomes.
- 3. Trisomy is the gain of a single chromosome, represented as 2n + 1. A human trisomic zygote has 47 chromosomes. The gain of a chromosome means that there are three homologous copies of one chromosome. Most cases of Down syndrome, discussed later in this chapter, result from trisomy of chromosome 21.
- 4. Tetrasomy is the gain of two homologous chromosomes, represented as 2n + 2. A human tetrasomic zygote has 48 chromosomes. Tetrasomy is not the gain of any two extra chromosomes, but rather the gain of two homologous chromosomes, so there will be four homologous copies of a particular chromosome.
223
More than one aneuploid mutation may occur in the same individual organism. An individual that has an extra copy of two different (nonhomologous) chromosomes is referred to as being double trisomic and represented as 2n + 1 + 1. Similarly, a double monosomic has two fewer nonhomologous chromosomes (2n − 1 − 1), and a double tetrasomic has two extra pairs of homologous chromosomes (2n + 2 + 2).
Effects of Aneuploidy
One of the first aneuploids to be recognized was a fruit fly with a single X chromosome and no Y chromosome discovered by Calvin Bridges in 1913. Another early study of aneuploidy focused on mutants in the Jimson weed, Datura stramonium. A. Francis Blakeslee began breeding this plant in 1913, and he observed that crosses with several Jimson mutants produced unusual ratios of progeny. For example, the globe mutant (producing a globe-shaped seedcase) was dominant but was inherited primarily from the female parent. When plants having the globe mutation were self-fertilized, only 25% of the progeny had the globe phenotype. If the globe mutant were strictly dominant, Blakeslee should have seen 75% of the progeny with the trait (see Chapter 3), and so the 25% that he observed was unsual. Blakeslee isolated 12 different mutants (Figure 8.20) that exhibited peculiar patterns of inheritance. Eventually, John Belling demonstrated that these 12 mutants are in fact trisomics. Datura stramonium has 12 pairs of chromosomes (2n = 24), and each of the 12 mutants is trisomic for a different chromosome pair. The aneuploid nature of the mutants explained the unusual ratios that Blakeslee had observed in the progeny. Many of the extra chromosomes in the trisomics were lost in meiosis, so fewer than 50% of the gametes carried the extra chromosome and the proportion of trisomics in the progeny was low. Furthermore, the pollen containing an extra chromosome was not as successful in fertilization, and trisomic zygotes were less viable.
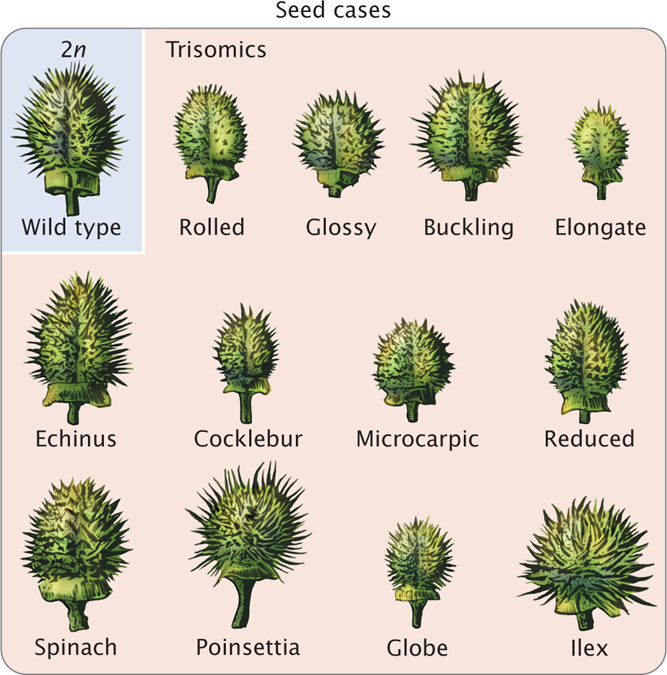
224
Aneuploidy usually alters the phenotype drastically. In most animals and many plants, aneuploid mutations are lethal. Because aneuploidy affects the number of gene copies but not their nucleotide sequences, the effects of aneuploidy are most likely due to abnormal gene dosage. Aneuploidy alters the dosage for some, but not all, genes, disrupting the relative concentrations of gene products and often interfering with normal development.
A major exception to the relation between gene number and gene dosage pertains to genes on the mammalian X chromosome. In mammals, X-chromosome inactivation ensures that males (who have a single X chromosome) and females (who have two X chromosomes) receive the same functional dosage for X-linked genes (see pp. 92–94 in Chapter 4 for further discussion of X-chromosome inactivation). Additional X chromosomes in mammals are inactivated, so we might expect that aneuploidy of the sex chromosomes would be less detrimental in these animals. Indeed, this is the case for mice and humans, for whom aneuploids of the sex chromosomes are the most common form of aneuploidy seen in living organisms. Y-chromosome aneuploids are probably common because there is so little information in the Y-chromosome.
CONCEPTS
Aneuploidy, the loss or gain of one or more individual chromosomes, may arise from the loss of a chromosome subsequent to translocation or from nondisjunction in meiosis or mitosis. It disrupts gene dosage and often has severe phenotypic effects.
 CONCEPT CHECK 5
CONCEPT CHECK 5
A diploid organism has 2n = 36 chromosomes. How many chromosomes will be found in a trisomic member of this species?
Aneuploidy in Humans
For unknown reasons, a high percentage of all human embryos that are conceived possess chromosome abnormalities. Findings from studies of women who are attempting pregnancy suggest that more than 30% of all conceptions spontaneously abort (miscarry), usually so early in development that the woman is not even aware of her pregnancy. Chromosome defects are present in at least 50% of spontaneously aborted human fetuses, with aneuploidy accounting for most of them. This rate of chromosome abnormality in humans is higher than in other organisms that have been studied; in mice, for example, aneuploidy is found in no more than 2% of fertilized eggs. Aneuploidy in humans usually produces such serious developmental problems that spontaneous abortion results. Only about 2% of all fetuses with a chromosome defect survive to birth.
Sex-Chromosome Aneuploids
The most common aneuploidies seen in living humans are those that involve the sex chromosomes. As is true of all mammals, aneuploidy of the human sex chromosomes is better tolerated than aneuploidy of autosomal chromosomes. Both Turner syndrome and Klinefelter syndrome (see Figures 4.8 and 4.9) result from aneuploidy of the sex chromosomes.
Autosomal Aneuploids
Autosomal aneuploids resulting in live births are less common than sex-chromosome aneuploids in humans, probably because there is no mechanism of dosage compensation for autosomal chromosomes. Most autosomal aneuploids are spontaneously aborted, though occasionally aneuploids of some of the small autosomes such as chromosome 21 complete development and result in a person with aneuploidy. Because these chromosomes are small and carry fewer genes, the presence of extra copies is less detrimental than it is for larger chromosomes.
Down Syndrome
In 1866, John Langdon Down, physician and medical superintendent of the Earlswood Asylum in Surrey, England, noticed a remarkable resemblance among a number of his intellectually disabled patients: all of them possessed a broad, flat face, a small nose, and oval-shaped eyes. Their features were so similar, in fact, that he felt that they might easily be mistaken as children of the same family. Down did not understand the cause of their intellectual disability, but his original description faithfully records the physical characteristics of this genetic form of intellectual disability. In his honor, the disorder is today known as Down syndrome.
225
Down syndrome, also known as trisomy 21, is the most common autosomal aneuploidy in humans (Figure 8.21a). The incidence of Down syndrome in the United States is similar to that in the world—about 1 in 700 human births—although the incidence increases among children born to older mothers. Approximately 92% of those who have Down syndrome have three full copies of chromosome 21 (and therefore a total of 47 chromosomes), a condition termed primary Down syndrome (Figure 8.21b). Primary Down syndrome usually arises from spontaneous nondisjunction in egg formation: about 75% of the nondisjunction events that cause Down syndrome are maternal in origin, most arising in meiosis I. Most children with Down syndrome are born to normal parents, and the failure of the chromosomes to divide has little hereditary tendency. A couple who has conceived one child with primary Down syndrome has only a slightly higher risk of conceiving a second child with Down syndrome (compared with other couples of similar age who have not had any Down-syndrome children). Similarly, the couple’s relatives are not more likely to have a child with primary Down syndrome.
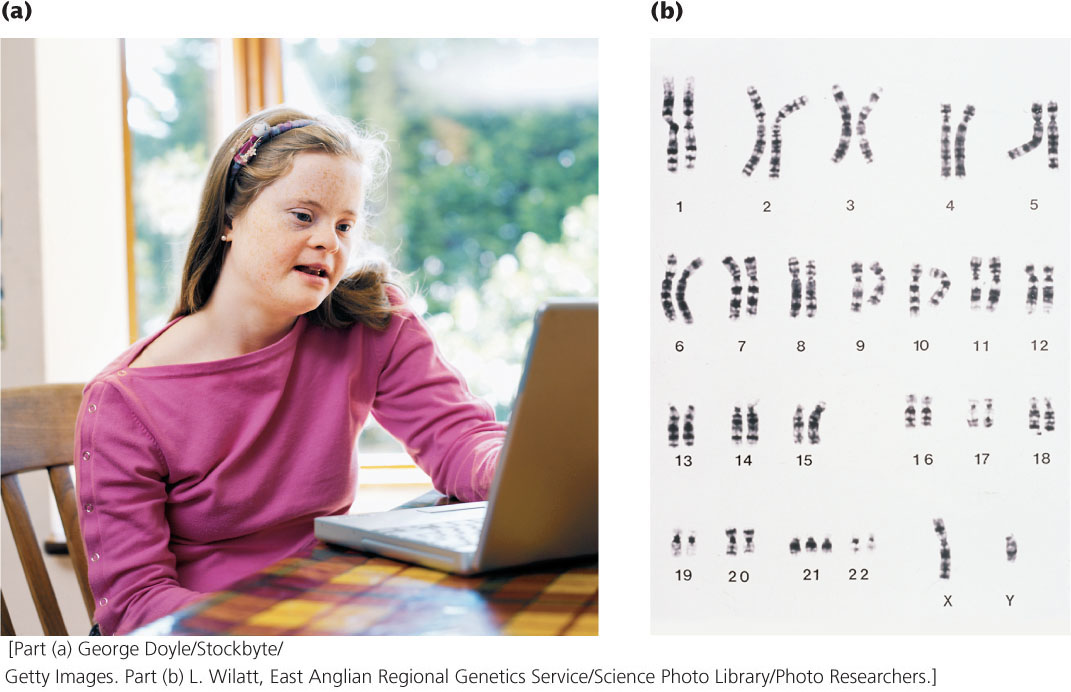
About 4% of people with Down syndrome are not trisomic for a complete chromosome 21. Instead, they have 46 chromosomes, but an extra copy of part of chromosome 21 is attached to another chromosome through a translocation. This condition is termed familial Down syndrome because it has a tendency to run in families. The phenotypic characteristics of familial Down syndrome are the same as those of primary Down syndrome.
Familial Down syndrome arises in offspring whose parents are carriers of chromosomes that have undergone a Robertsonian translocation, most commonly between chromosome 21 and chromosome 14: the long arm of 21 and the short arm of 14 exchange places (Figure 8.22). This exchange produces a chromosome that includes the long arms of chromosomes 14 and 21, and a very small chromosome that consists of the short arms of chromosomes 21 and 14. The small chromosome is generally lost after several cell divisions. Although exchange between chromosomes 21 and 14 is the most common cause of familial Down syndrome, the condition can also be caused by translocations between 21 and other chromosomes such as 15.
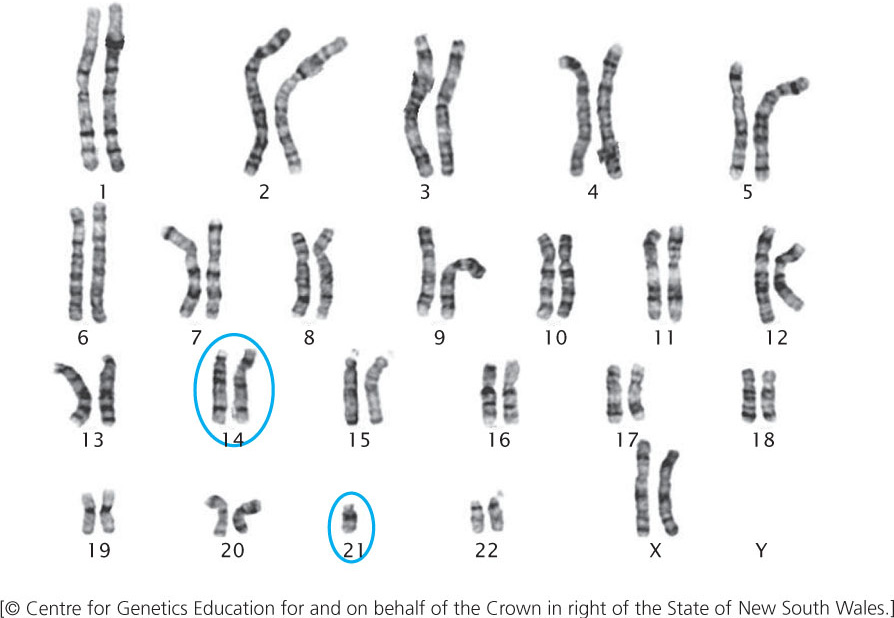
226
Persons with the translocation, called translocation carriers, do not have Down syndrome. Although they possess only 45 chromosomes, their phenotypes are normal because they have two copies of the long arms of chromosomes 14 and 21, and apparently the short arms of these chromosomes (which are lost) carry no essential genetic information. Although translocation carriers are completely healthy, they have an increased chance of producing children with Down syndrome.
When a translocation carrier produces gametes, the translocation chromosome may segregate in three different ways. First, it may separate from the normal chromosomes 14 and 21 in anaphase I of meiosis (Figure 8.23a). In this type of segregation, half of the gametes will have the translocation chromosome and no other copies of chromosomes 21 and 14; the fusion of such a gamete with a normal gamete will give rise to a translocation carrier. The other half of the gametes produced by this first type of segregation will be normal, each with a single copy of chromosomes 21 and 14, and will result in normal offspring.
Alternatively, the translocation chromosome may separate from chromosome 14 and pass into the same cell with the normal chromosome 21 (Figure 8.23b). This type of segregation produces abnormal gametes only; half will have two functional copies of chromosome 21 (one normal and one attached to chromosome 14) and the other half will lack chromosome 21. If a gamete with the two functional copies of chromosome 21 fuses with a normal gamete carrying a single copy of chromosome 21, the resulting zygote will have familial Down syndrome. If a gamete lacking chromosome 21 fuses with a normal gamete, the resulting zygote will have monosomy 21 and will be spontaneously aborted.
In the third type of segregation, the translocation chromosome and the normal copy of chromosome 14 segregate together (Figure 8.23c). This pattern is presumably rare, because the two centromeres are both derived from chromosome 14 and usually separate from each other. All the gametes produced by this process are abnormal: half result in monosomy 14 and the other half result in trisomy 14. All are spontaneously aborted.
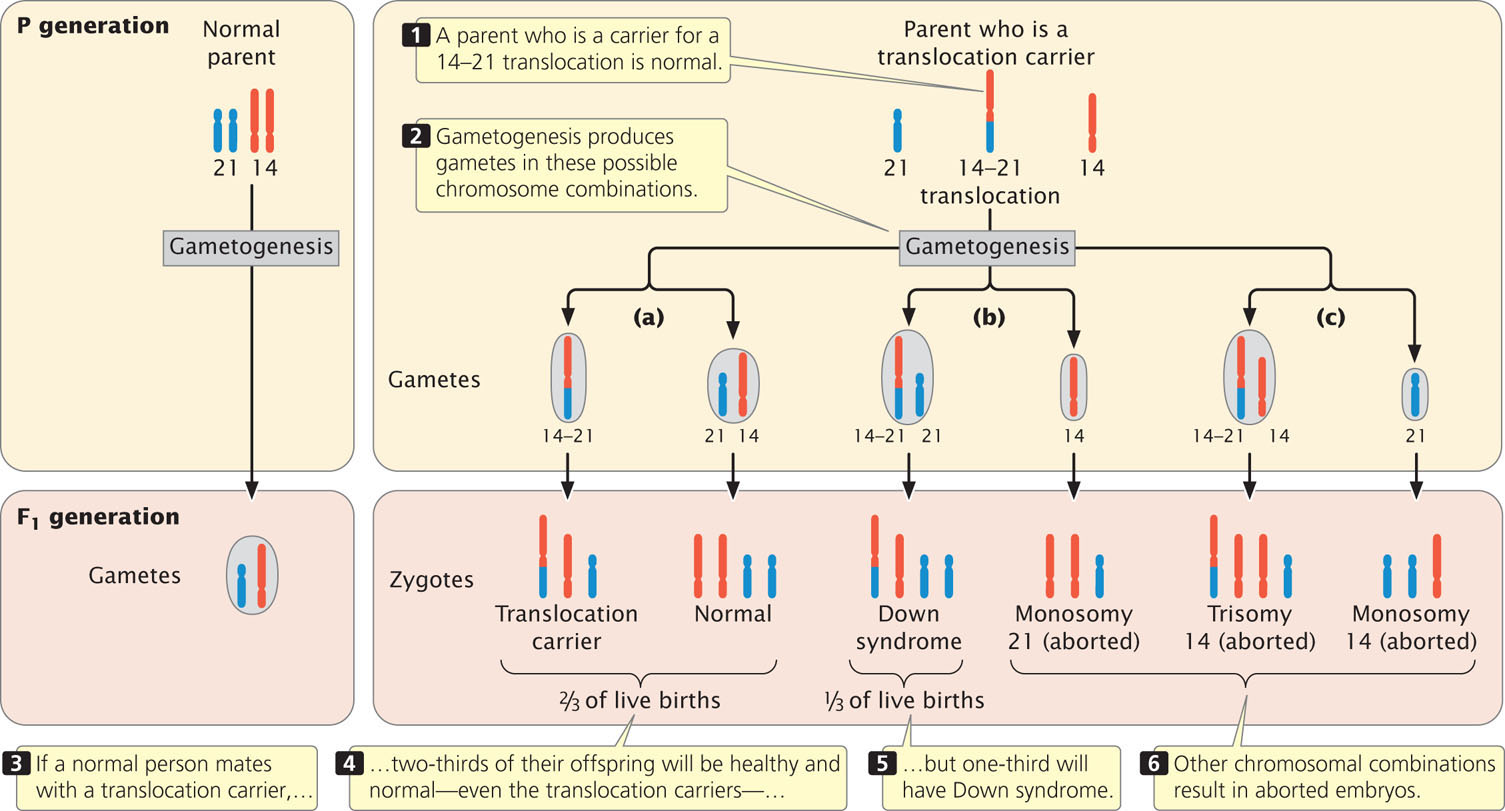
Thus, only three of the six types of gametes that can be produced by a translocation carrier will result in the birth of a baby and, theoretically, these gametes should arise with equal frequency. One-third of the offspring of a translocation carrier should be translocation carriers like their parent, one-third should have familial Down syndrome, and one-third should be normal. In reality, however, fewer than one-third of the children born to translocation carriers have Down syndrome, which suggests that some of the embryos with Down syndrome are spontaneously aborted. ![]() TRY PROBLEM 32
TRY PROBLEM 32
227
Other Human Trisomies
Few autosomal aneuploids in humans besides trisomy 21 result in live births. Trisomy 18, also known as Edward syndrome, arises with a frequency of approximately 1 in 8000 live births. Babies with Edward syndrome have severe intellectual disability, low-set ears, a short neck, deformed feet, clenched fingers, heart problems, and other disabilities. Few live for more than a year after birth. Trisomy 13 has a frequency of about 1 in 15,000 live births and produces features that are collectively known as Patau syndrome. Characteristics of this condition include severe intellectual disability, a small head, sloping forehead, small eyes, cleft lip and palate, extra fingers and toes, and numerous other problems. About half of children with trisomy 13 die within the first month of life, and 95% die by the age of 3. Rarer still is trisomy 8, which arises with a frequency ranging from about 1 in 25,000 to 1 in 50,000 live births. This aneuploid is characterized by intellectual disability, contracted fingers and toes, low-set malformed ears, and a prominent forehead. Many who have this condition have normal life expectancy.
Aneuploidy and Maternal Age
Most cases of Down syndrome and other types of aneuploidy in humans arise from maternal nondisjunction, and the frequency of aneuploidy increases with maternal age (Figure 8.24). Why maternal age is associated with nondisjunction is not known for certain. Female mammals are born with primary oocytes suspended in the diplotene substage of prophase I of meiosis. Just before ovulation, meiosis resumes and the first division is completed, producing a secondary oocyte. At this point, meiosis is suspended again and remains so until the secondary oocyte is penetrated by a sperm. The second meiotic division takes place immediately before the nuclei of egg and sperm unite to form a zygote.
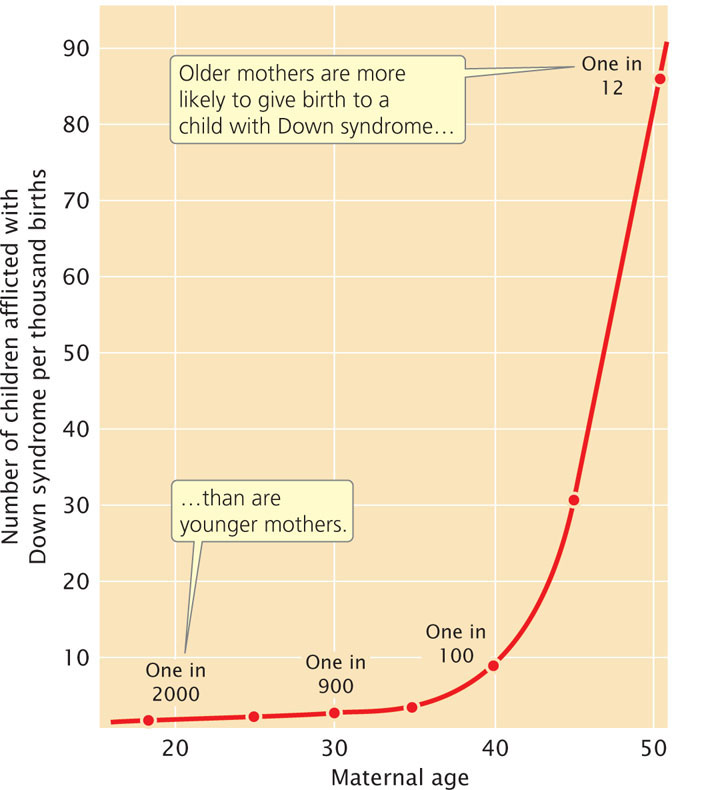
Primary oocytes may remain suspended in diplotene for many years before ovulation takes place and meiosis recommences. Components of the spindle and other structures required for chromosome segregation may break down in the long arrest of meiosis, leading to more aneuploidy in children born to older mothers. According to this theory, no age effect is seen in males, because sperm are produced continuously after puberty with no long suspension of the meiotic divisions.
Aneuploidy and Cancer
Many tumor cells have extra or missing chromosomes, or both; some types of tumors are consistently associated with specific chromosome mutations, including aneuploidy and chromosome rearrangements. The role of chromosome mutations in cancer will be explored in Chapter 23.
CONCEPTS
In humans, sex-chromosome aneuploids are more common than autosomal aneuploids. X-chromosome inactivation prevents problems of gene dosage for X-linked genes. Down syndrome results from three functional copies of chromosome 21, either through trisomy (primary Down syndrome) or a Robertsonian translocation (familial Down syndrome).
CONCEPT CHECK 6
Briefly explain why, in humans and mammals, sex-chromosome aneuploids are more common than autosomal aneuploids.
Uniparental Disomy
Normally, the two chromosomes of a homologous pair are inherited from different parents—one from the father and one from the mother. The development of molecular techniques that facilitate the identification of specific DNA sequences (see Chapter 19) has made the determination of the parental origins of chromosomes possible. Surprisingly, sometimes both chromosomes are inherited from the same parent, a condition termed uniparental disomy.
Many cases of uniparental disomy probably originate as a trisomy. Although most autosomal trisomies are lethal, a trisomic embryo can survive if one of the three chromosomes is lost early in development. If, just by chance, the two remaining chromosomes are both from the same parent, uniparental disomy results.
Uniparental disomy violates the rule that children affected with a recessive disorder appear only in families where both parents are carriers. For example, cystic fibrosis is an autosomal recessive disease; typically, both parents of an affected child are heterozygous for the cystic fibrosis mutation on chromosome 7. However, for a small proportion of people with cystic fibrosis, only one of the parents is heterozygous for the cystic fibrosis mutation. In these cases, cystic fibrosis is due to uniparental disomy: the person who has cystic fibrosis inherited from the heterozygous parent two copies of the chromosome 7 that carries the defective cystic fibrosis allele and no copy of the normal allele from the other parent.
228
Uniparental disomy has also been observed in Prader–Willi syndrome, a rare condition that arises when a paternal copy of a gene on chromosome 15 is missing. Although most cases of Prader–Willi syndrome result from a chromosome deletion that removes the paternal copy of the gene, 20% to 30% of the cases arise when both copies of chromosome 15 are inherited from the mother and no copy is inherited from the father.
Mosaicism
Nondisjunction in a mitotic division may generate patches of cells in which every cell has a chromosome abnormality and other patches in which every cell has a normal karyotype. This type of nondisjunction leads to regions of tissue with different chromosome constitutions, a condition known as mosaicism. Growing evidence suggests that mosaicism is more common that is often recognized. For example, about 50% of those diagnosed with Turner syndrome (individuals with a single X chromosome) are actually mosaics, possessing some 45,X cells and some normal 46,XX cells. A few may even be mosaics for two or more types of abnormal karyotypes. The 45,X/46,XX mosaic usually arises when an X chromosome is lost soon after fertilization in an XX embryo.
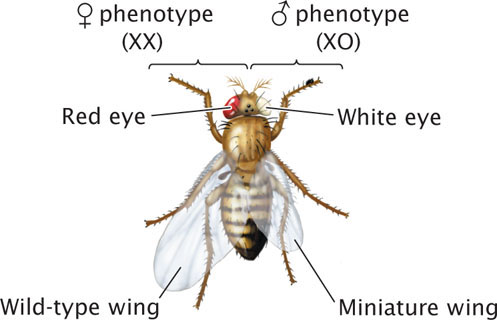
Fruit flies that are XX/XO mosaics (O designates the absence of a homologous chromosome; XO means that the cell has a single X chromosome and no Y chromosome) develop a mixture of male and female traits, because the presence of two X chromosomes in fruit flies produces female traits and the presence of a single X chromosome produces male traits (Figure 8.25). In fruit flies, sex is determined independently in each cell in the course of development. Those cells that are XX express female traits; those that are XO express male traits. Such sexual mosaics are called gynandromorphs. Normally, X-linked recessive genes are masked in heterozygous females but, in XX/XO mosaics, any X-linked recessive genes present in the cells with a single X chromosome will be expressed.
CONCEPTS
In uniparental disomy, an individual organism has two copies of a chromosome from one parent and no copy from the other. Uniparental disomy may arise when a trisomic embryo loses one of the triplicate chromosomes early in development. In mosaicism, different cells within the same individual organism have different chromosome constitutions.