Positive Control and Catabolite Repression
Escherichia coli and many other bacteria metabolize glucose preferentially in the presence of lactose and other sugars. They do so because glucose enters glycolysis without further modification and therefore requires less energy to metabolize than do other sugars. When glucose is available, genes that participate in the metabolism of other sugars are repressed through a process known as catabolite repression. Efficient transcription of the lac operon, for example, takes place only if lactose is present and glucose is absent. But how is the expression of the lac operon influenced by glucose? What brings about catabolite repression?
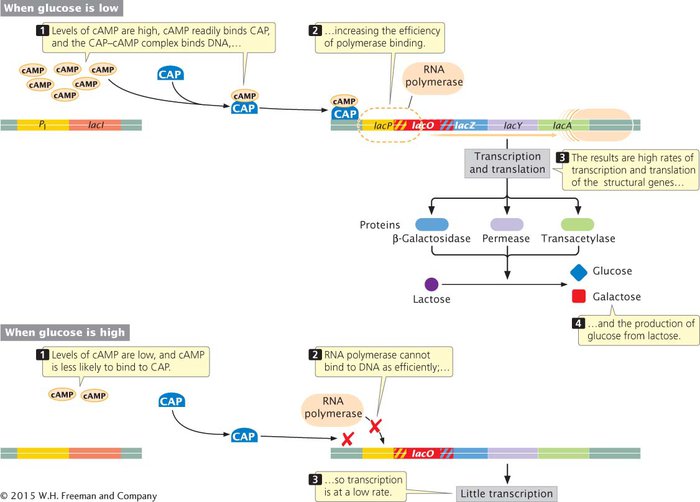
Catabolite repression results from positive control in response to glucose. (This regulation is in addition to the negative control brought about by repressor binding to the operator of the lac operon when lactose is absent.) Positive control is accomplished through the binding of a protein called the catabolite activator protein (CAP) to a site about 22 nucleotides long that is located within or slightly upstream of the promoter of the lac genes (Figure 12.12). RNA polymerase does not bind efficiently to some promoters unless CAP is first bound to the DNA. Before CAP can bind to DNA, however, it must form a complex with a modified nucleotide called adenosine- TRY PROBLEM 20
TRY PROBLEM 20
CONCEPTS
In spite of its name, catabolite repression is a type of positive control of the lac operon. The catabolite activator protein (CAP), complexed with cAMP, binds to a site near the promoter and stimulates the binding of RNA polymerase. Cellular levels of cAMP in the cell are controlled by glucose; a low glucose level increases the abundance of cAMP and enhances the transcription of the lac structural genes.