Natural Selection
A final process that brings about changes in allelic frequencies is natural selection, the differential reproduction of genotypes (see Section 17.4). Natural selection takes place when individuals with adaptive traits produce a greater number of offspring than that produced by others in the population. If the adaptive traits have a genetic basis, they are inherited by the offspring and appear with greater frequency in the next generation. A trait that provides a reproductive advantage thereby increases over time, enabling populations to become better suited—

FITNESS AND THE SELECTION COEFFICIENT The effect of natural selection on the gene pool of a population depends on the fitness values of the genotypes in the population. Fitness is defined as the relative reproductive success of a genotype. Here, the term relative is critical: fitness is the reproductive success of one genotype compared with the reproductive successes of other genotypes in the population.
Fitness values range from 0 to 1. Suppose that the average number of viable offspring produced by three genotypes is
| Genotypes: | A1A1 | A1A2 | A2A2 |
|---|---|---|---|
| Mean number of offspring produced: | 10 | 5 | 2 |
To calculate fitness (W) for each genotype, we take the mean number of offspring produced by a genotype and divide it by the mean number of offspring produced by the most prolific genotype:
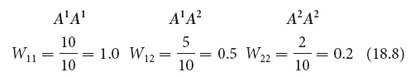
The fitness of genotype A1A1 is designated W11, that of A1A2 is W12, and that of A2A2 is W22.
A related variable is the selection coefficient (s), which is the relative intensity of selection against a genotype. We usually speak of selection for a particular genotype, but keep in mind that, when selection is for one genotype, selection is automatically against at least one other genotype. The selection coefficient is equal to 1 − W; so the selection coefficients for the preceding three genotypes are
A1A1 A1A2 A2A2
s11 = 0 s12 = 0.5 s22 = 0.8
CONCEPTS
Natural selection is the differential reproduction of genotypes. It is measured as fitness, which is the reproductive success of a genotype compared with that of other genotypes in a population.
 CONCEPT CHECK 5
CONCEPT CHECK 5
The average numbers of offspring produced by three genotypes are GG = 6; Gg = 3, gg = 2. What is the fitness of Gg?
3
0.5
0.3
0.27
b
THE RESULTS OF SELECTION The results of selection depend on the fitnesses of the genotypes in a population. In a population with three genotypes (A1A1, A1A2, and A2A2) with fitnesses W11, W12, and W22, we can identify six different types of natural selection (Table 18.1).
| Type | Fitness relation | Form of selection | Result |
|---|---|---|---|
| 1 | W11 = W12 > W22 | Directional selection against recessive allele A2 | A1 increases, A2 decreases |
| 2 | W11 = W12 < W22 | Directional selection against dominant allele A1 | A2 increases, A1 decreases |
| 3 | W11 > W12 > W22 | Directional selection against incompletely dominant allele A2 | A1 increases, A2 decreases |
| 4 | W11 < W12 < W22 | Directional selection against incompletely dominant allele A1 | A2 increases, A1 decreases |
| 5 | W11 < W12 > W22 | Overdominance | Stable equilibrium, both alleles maintained |
| 6 | W11 > W12 < W22 | Underdominance | Unstable equilibrium |
|
Note: W11, W12, and W22 represent the fitnesses of genotypes A1A1, A1A2, and A2A2, respectively. |
|||
In type 1 selection, a dominant allele A1 confers a fitness advantage; in this case, the fitnesses of genotypes A1A1 and A1A2 are equal and higher than the fitness of A2A2 (W11 = W12 > W22). Because both the heterozygote and the A1A1 homozygote have copies of the A1 allele and produce more offspring than the A2A2 homozygote does, the frequency of the A1 allele will increase with time, and the frequency of the A2 allele will decrease. This form of selection, in which one allele or trait is favored over another, is termed directional selection.
Type 2 selection is directional selection against a dominant allele A1 (W11 = W12 < W22). Type 3 and type 4 selection are also directional selection, but in these cases, there is incomplete dominance, and the heterozygote has a fitness that is intermediate between the two homozygotes (W11 > W12 > W22 for type 3; W11 < W12 < W22 for type 4). Eventually, all four types of directional selection lead to fixation of the favored allele and elimination of the other allele, as long as no other evolutionary forces act on the population.
The last two types of selection (types 5 and 6) are special situations that lead to equilibrium, where there is no further change in allelic frequency. Type 5 selection is referred to as overdominance or heterozygote advantage. Here, the heterozygote has higher fitness than either homozygote (W11 < W12 > W22). With overdominance, both alleles are favored in the heterozygote, and neither allele is eliminated from the population. The allelic frequencies change with overdominant selection until a stable equilibrium is reached, at which point there is no further change. The allelic frequency at equilibrium (^q) depends on the fitnesses (usually expressed as selection coefficients) of the two homozygotes:
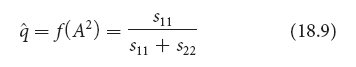
where s11 represents the selection coefficient of the A1A1 homozygote and s22 represents the selection coefficient of the A2A2 homozygote.
An example of overdominance is sickle-
The last type of selection (type 6) is underdominance, in which the heterozygote has lower fitness than either homozygote (W11 > W12 < W22). Underdominance leads to an unstable equilibrium; here, allelic frequencies do not change as long as they are at equilibrium, but if they are disturbed from the equilibrium point by some other evolutionary force, they will move away from equilibrium until one allele eventually becomes fixed.
CONCEPTS
Natural selection changes allelic frequencies; the direction and magnitude of change depend on the intensity of selection, the dominance relations of the alleles, and the allelic frequencies. Directional selection favors one allele over another and eventually leads to fixation of the favored allele. Overdominance leads to a stable equilibrium with maintenance of both alleles in the population. Underdominance produces an unstable equilibrium because the heterozygote has lower fitness than either homozygote.
CONNECTING CONCEPTS
The General Effects of Forces That Change Allelic Frequencies
You now know that four processes can bring about change in the allelic frequencies of a population: mutation, migration, genetic drift, and natural selection. Their short-
| Force | Short- |
Long- |
|---|---|---|
| Mutation | Change in allelic frequencies | Equilibrium reached between forward and reverse mutations |
| Migration | Change in allelic frequencies | Equilibrium reached when allelic frequencies of source and recipient population are equal |
| Genetic drift | Change in allelic frequencies | Fixation of one allele |
| Natural selection | Change in allelic frequencies | Directional selection: fixation of one allele Overdominant selection: equilibrium reached Underdominant selection: unstable equilibrium |
These evolutionary forces affect both genetic variation within populations and genetic divergence between populations. Evolutionary forces that maintain or increase genetic variation within populations are listed in the upper left quadrant of Figure 18.8. These forces include some types of natural selection, such as overdominance, in which both alleles are favored. Mutation and migration also increase genetic variation within populations because they introduce new alleles to a population. Evolutionary forces that decrease genetic variation within populations are listed in the lower left quadrant of Figure 18.8. These forces include genetic drift, which decreases variation through the fixation of alleles, and some forms of natural selection, such as directional selection.
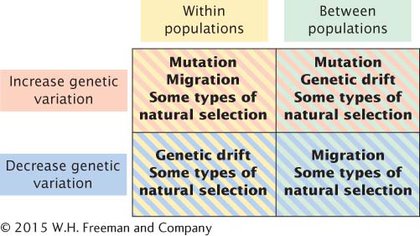
These same evolutionary forces also affect genetic divergence between populations. Natural selection increases divergence between populations if different alleles are favored in different populations, but it can also decrease divergence between populations by favoring the same allele in different populations. Mutation almost always increases divergence between populations because different mutations arise in each population. Genetic drift also increases divergence between populations because changes in allelic frequencies due to drift are random and are likely to occur in different directions in separate populations. Migration, on the other hand, decreases divergence between populations because it makes populations more similar in their genetic composition.
Migration and genetic drift act in opposite directions: migration increases genetic variation within populations and decreases divergence between populations, whereas genetic drift decreases genetic variation within populations and increases divergence between populations. Mutation increases both variation within populations and divergence between populations. Natural selection can either increase or decrease variation within populations, and it can increase or decrease divergence between populations.
An important point to keep in mind is that real populations are simultaneously affected by many evolutionary forces. We have examined the effects of mutation, migration, genetic drift, and natural selection in isolation so that the effect of each process will be clear. In the real world, however, populations are commonly affected by several evolutionary forces at the same time, and evolution results from the complex interplay of numerous processes.