Modes of Speciation
Speciation is the process by which new species arise. In the terms of the biological species concept, speciation comes about through the evolution of reproductive isolating mechanisms.
New species arise in two principal ways. Allopatric speciation occurs when a geographic barrier splits a population into two groups and blocks the exchange of genes between them. The interruption of gene flow then leads to the evolution of genetic differences that result in reproductive isolation. Sympatric speciation is speciation that arises in the absence of any external barrier to gene flow; reproductive isolating mechanisms evolve within a single population. We will take a more detailed look at each of these mechanisms next.
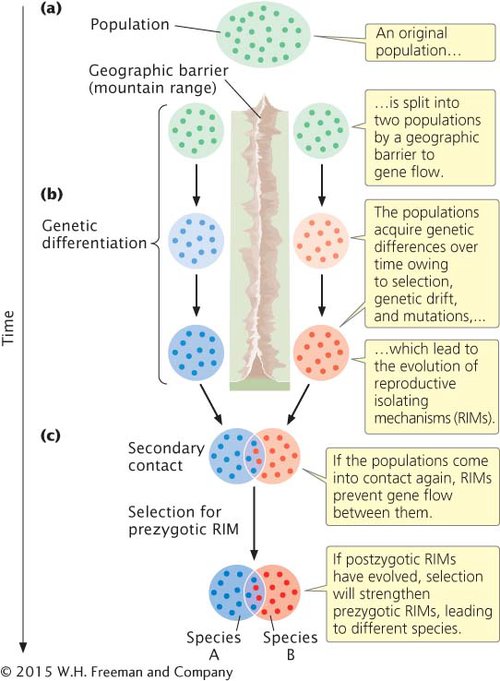
ALLOPATRIC SPECIATION Allopatric speciation is initiated when a geographic barrier splits a population into two or more groups and prevents gene flow between those groups (Figure 18.10a). Geographic barriers can take a number of forms. Uplifting of a mountain range may split a population of lowland plants into separate groups on each side of the mountains. Oceans serve as effective barriers for many types of terrestrial organisms, separating individuals on different islands from one another and from those on the mainland. Rivers often separate populations of fish located in separate drainages. The erosion of mountains may leave populations of alpine plants isolated on separate mountain peaks.
After two populations have been separated by a geographic barrier that prevents gene flow between them, they evolve independently (Figure 18.10b). The genetic isolation allows each population to accumulate genetic differences that are not found in the other population; genetic differences arise through natural selection, unique mutations, and genetic drift (if the populations are small). Genetic differentiation may eventually lead to prezygotic and postzygotic reproductive isolation. It is important to note that both types of reproductive isolation arise simply as a consequence of genetic divergence.
If the populations later come into secondary contact (Figure 18.10c), several outcomes are possible. If limited genetic differentiation has taken place during the separation of the populations, reproductive isolating mechanisms may not have evolved or may be incomplete. In this case, the populations will remain a single species. A second possible outcome is that genetic differentiation during separation has led to prezygotic reproductive isolating mechanisms; in this case, the two populations are different species. A third possible outcome is that, during their time apart, some genetic differentiation took place between the populations, leading to postzygotic reproductive isolation. If postzygotic isolating mechanisms have evolved, any mating between individuals from the different populations will produce hybrid offspring that are inviable or sterile. Individuals that mate only with members of the same population will have higher fitness than individuals that mate with members of the other population; therefore, natural selection will increase the frequency of any trait that prevents interbreeding between members of the different populations. Over time, prezygotic reproductive isolating mechanisms will evolve. In short, if some postzygotic reproductive isolation exists, natural selection will favor the evolution of prezygotic reproductive isolating mechanisms to prevent wasted reproductive effort by individuals mating with members of the other population. This process of postzygotic reproductive isolation leading to the evolution of prezygotic isolating mechanisms is termed reinforcement.
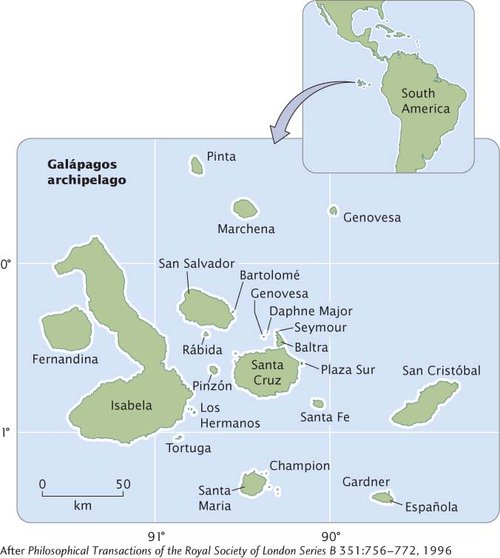
An excellent example of allopatric speciation can be found in Darwin’s finches, a group of birds that Charles Darwin discovered on the Galápagos Islands during his voyage aboard the Beagle. The Galápagos Islands form an archipelago located approximately 900 kilometers off the coast of South America (Figure 18.11). Consisting of more than a dozen large islands and many smaller ones, the Galápagos formed from volcanoes that erupted over a geological hot spot that has remained stationary while the geological plate over it moved eastward in the past 3 million years. The movement of the geological plate pulled newly formed islands eastward, so that the islands to the east (San Cristóbal and Española) are older than those to the west (Isabela and Fernandina). Over time, the number of islands in the archipelago increased as new volcanoes arose.
Darwin’s finches consist of 14 species found on various islands in the Galápagos archipelago. The birds vary in the shapes and sizes of their beaks, which are adapted for eating different types of food. Recent studies of the development of finch embryos have helped to reveal some of the molecular details of how differences in beak shapes have evolved. Genetic studies have demonstrated that all the birds are closely related and that they evolved from a single ancestral species that migrated to the islands from the coast of South America some 2 million to 3 million years ago. The evolutionary relationships among the 14 species, based on studies of microsatellite data (see Chapter 14), are depicted in Figure 18.12. Most of the species are separated by a behavioral isolating mechanism (song in particular), but some of the species can and occasionally do hybridize in nature.
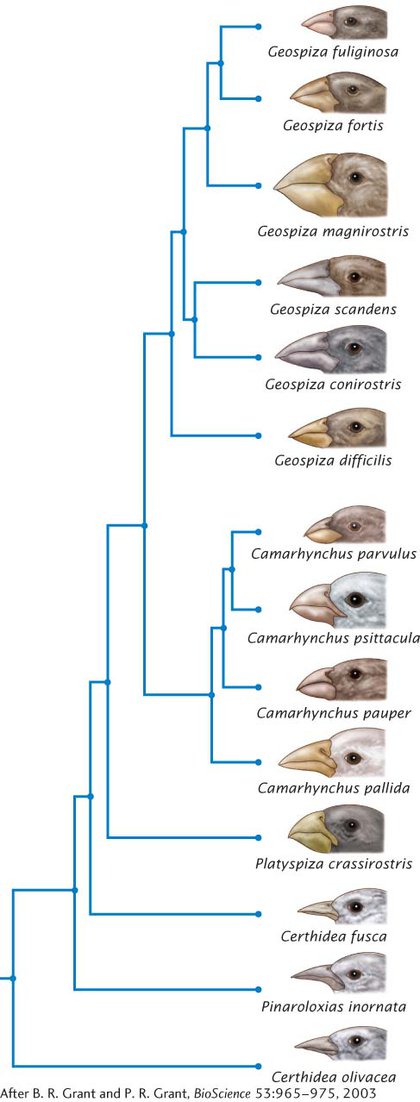
The first finches to arrive in the Galápagos probably colonized one of the larger eastern islands. A breeding population became established and increased with time. At some point, a few birds dispersed to another island, where they were effectively isolated from the original population, and established a new population. The new population underwent genetic differentiation owing to genetic drift and adaptation to the local conditions of the island, and it eventually became reproductively isolated from the original population. Individual birds from the new population then dispersed to other islands and gave rise to additional species. This process was repeated many times. Occasionally, newly evolved species dispersed to an island where another species was already present, giving rise to secondary contact between the species. Today, many of the islands have more than one resident finch species.
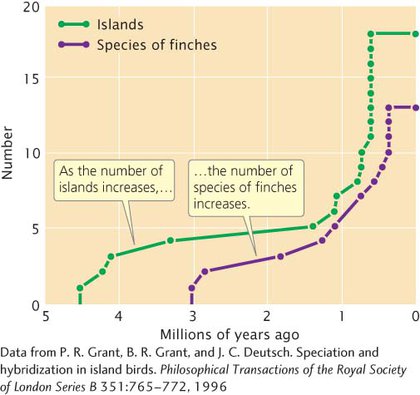
The ages of the 14 species have been estimated with data from mitochondrial DNA. As Figure 18.13 shows, there is a strong correspondence between the number of bird species present at various times in the past and the number of islands in the archipelago. This correspondence is one of the most compelling pieces of evidence for the theory that the different species of finches arose through allopatric speciation.
CONCEPTS
Allopatric speciation is initiated when a geographic barrier to gene flow splits a single population into two or more populations. Over time, the populations evolve genetic differences that bring about reproductive isolation. After postzygotic reproductive isolating mechanisms have evolved, selection favors the evolution of prezygotic reproductive isolating mechanisms.
 CONCEPT CHECK 7
CONCEPT CHECK 7
What role does genetic drift play in allopatric speciation?
Genetic drift can bring about changes in the allelic frequencies of populations and lead to genetic differences among populations. Genetic differentiation is the cause of postzygotic and prezygotic reproductive isolation between populations that leads to speciation.
SYMPATRIC SPECIATION Sympatric speciation arises in the absence of any geographic barrier to gene flow; reproductive isolating mechanisms evolve within a single interbreeding population. Sympatric speciation has long been a controversial topic within evolutionary biology. Ernst Mayr believed that sympatric speciation was impossible, and he demonstrated that many apparent cases of sympatric speciation could be explained by allopatric speciation. More recently, however, evidence has accumulated that sympatric speciation can arise, and has arisen, under special circumstances. The difficulty with sympatric speciation is that isolating mechanisms arise as a consequence of genetic differentiation, which takes place only if gene flow between groups is interrupted. But without reproductive isolation (or some external barrier), how can gene flow be interrupted? How can genetic differentiation arise within a single group that is freely exchanging genes?
Most models of sympatric speciation assume that genetic differentiation is initiated by strong selection favoring different phenotypes within a single population. An example of how sympatric speciation might arise is seen in the apple maggot flies (Rhagoletis pomonella) (Figure 18.14) studied by Guy Bush and others. The flies of this species feed on the fruits of a specific host tree. Mating takes place near the fruits, and the flies lay their eggs on the ripened fruits, where their larvae grow and develop. Rhagoletis pomonella originally lived only on the fruits of hawthorn trees, which are native to North America. One hundred fifty years ago, R. pomonella was first observed on cultivated apples, which are related to hawthorns but a different species. Infestations of apples by this new apple host race of R. pomonella quickly spread, and today, many apple trees throughout North America are infested with the flies.

The apple host race of R. pomonella probably originated when a few flies acquired a mutation that allowed them to feed on apples instead of hawthorn fruits. Because mating takes place on and near fruits, flies that use apples are likely to mate with other flies that use apples, leading to genetic isolation between flies using hawthorns and those using apples. Indeed, Bush found that some genetic differentiation has already taken place between the two host races. Furthermore, the flies lay their eggs on ripening fruit, so there has been strong selection on the flies to synchronize their reproduction with the period when their host species has ripening fruit. Apples ripen several weeks earlier than hawthorns do. Correspondingly, the peak mating period of the apple host race is 3 weeks earlier than that of the hawthorn host race. These differences in the timing of reproduction between apple and hawthorn host races have further reduced gene flow—