Transduction: Using Phages To Map Bacterial Genes
In our discussion of bacterial genetics, three mechanisms of gene transfer were identified: conjugation, transformation, and transduction (see Figure 7.7). Let’s take a closer look at transduction, in which genes are transferred between bacteria by viruses. In generalized transduction, any gene may be transferred. In specialized transduction, which will not be discussed in detail here, only a few genes are transferred.
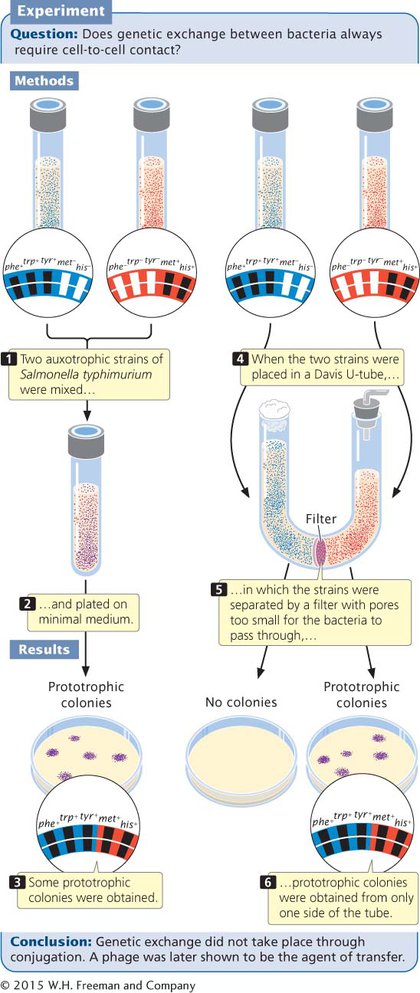
Joshua Lederberg and Norton Zinder discovered generalized transduction in 1952 while trying to produce recombination in the bacterium Salmonella typhimurium by conjugation. They mixed a strain of S. typhimurium that was phe+ trp+ tyr+ met− his− with a strain that was phe− trp− tyr− met+ his+ (Figure 7.22) and plated the mixture on minimal medium. A few prototrophic recombinants (phe+ trp+ tyr+ met+ his+) appeared, suggesting that conjugation had taken place. However, when they tested the two strains in a U-
In the lytic cycle of phage reproduction, the phage degrades the bacterial chromosome into random fragments (Figure 7.23). In some types of bacteriophage, a piece of the bacterial chromosome, instead of phage DNA, occasionally gets packaged into a phage coat; these phage particles are called transducing phages. If the transducing phage infects a new cell and releases the bacterial DNA, the introduced genes may then become integrated into the bacterial chromosome by a double crossover. Some transducing phages insert viral DNA, along with the bacterial gene, into the bacterial chromosome. In either case, bacterial genes can be moved from one bacterial strain to another, producing recombinant bacteria called transductants.
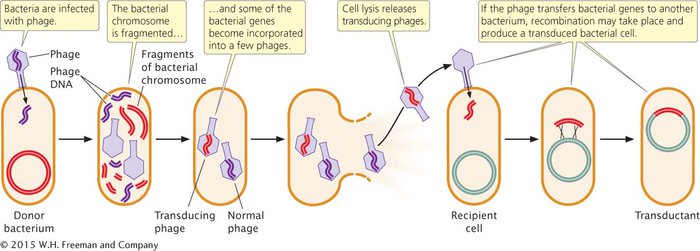
Not all phages are capable of transduction, a rare event that requires (1) that the phage degrade the bacterial chromosome, (2) that the process of packaging DNA into the phage particle not be specific for phage DNA, and (3) that the bacterial genes transferred by the virus recombine with the chromosome in the recipient cell. The overall rate of transduction ranges from about 1 in 100,000 to 1 in 1,000,000 phages.
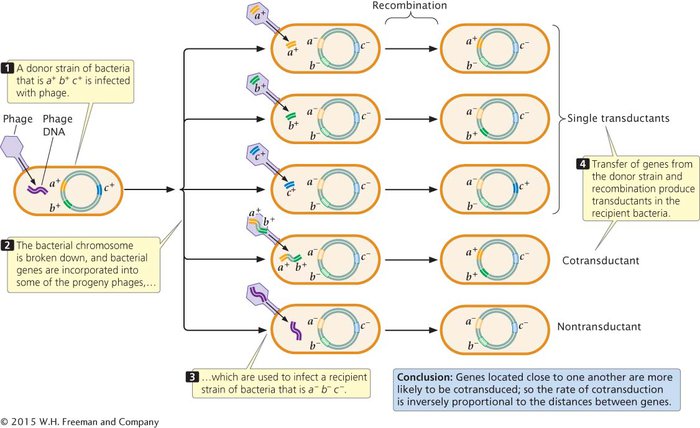
Because of the limited size of a phage particle, only about 1% of the bacterial chromosome can be transduced. Only genes located close together on the bacterial chromosome will be transferred together, or cotransduced. Because the chance of a cell undergoing transduction by two separate phages is exceedingly small, we can assume that any cotransduced genes are located close together on the bacterial chromosome. Thus, rates of cotransduction, like rates of cotransformation, give an indication of the physical distances between genes on a bacterial chromosome.
To map genes by using transduction, two bacterial strains with different alleles at several loci are used. The donor strain is infected with phages (Figure 7.24), which reproduce within the cells. When the phages have lysed the donor cells, a suspension of the progeny phages is mixed with a recipient strain of bacteria, which is then plated on several different kinds of media to determine the phenotypes of the transducing progeny phages.  TRY PROBLEM 20
TRY PROBLEM 20
CONCEPTS
In transduction, bacterial genes become packaged into a viral coat, are transferred to another bacterium by the virus, and become incorporated into the bacterial chromosome by crossing over. Bacterial genes can be mapped with the use of generalized transduction.
 CONCEPT CHECK 6
CONCEPT CHECK 6
In gene mapping using generalized transduction, bacterial genes that are cotransduced are
far apart on the bacterial chromosome.
on different bacterial chromosomes.
close together on the bacterial chromosome.
on a plasmid.
c
CONNECTING CONCEPTS
Three Methods for Mapping Bacterial Genes
Three methods of mapping bacterial genes have now been outlined: (1) interrupted conjugation mapping, (2) transformation mapping, and (3) transduction mapping. These methods have important similarities and differences.
Mapping with interrupted conjugation is based on the time required for genes to be transferred from one bacterium to another by means of cell-
In gene mapping with transformation, DNA from the donor strain is isolated, broken up, and mixed with the recipient strain. Some fragments pass into the recipient cells, where the transformed DNA may recombine with the bacterial chromosome. The unit of transfer here is a random fragment of the chromosome. Loci that are close together on the donor chromosome tend to be on the same DNA fragment, so the rates of cotransformation provide information about the relative positions of genes on the chromosome.
Transduction mapping also relies on the transfer of genes between bacteria that differ in two or more traits, but here, the vehicle of gene transfer is a bacteriophage. In a number of respects, transduction mapping is similar to transformation mapping. Small fragments of DNA are carried by the phage from donor to recipient bacteria, and the rates of cotransduction, like the rates of cotransformation, provide information about the relative distances between the genes.
All of the methods use a common strategy for mapping bacterial genes. The movement of genes from donor to recipient is detected by using strains that differ in two or more traits, and the transfer of one gene relative to the transfer of others is examined. Additionally, all three methods rely on recombination between the transferred DNA and the bacterial chromosome. In mapping with interrupted conjugation, the relative order and timing of gene transfer provide the information necessary to map the genes; in transformation and transduction mapping, the rate of cotransfer provides this information.
In conclusion, the same basic strategies are used for mapping with interrupted conjugation, transformation, and transduction. The methods differ principally in their mechanisms of transfer: in interrupted conjugation mapping, DNA is transferred though contact between bacteria; in transformation mapping, DNA is transferred as small naked fragments; and, in transduction mapping, DNA is transferred by bacteriophages.