CONCEPT4.1 Cells Provide Compartments for Biochemical Reactions
Cells contain water and other small and large molecules, which we examined in Chapters 2 and 3. Each cell contains at least 10,000 different types of molecules, most of them present in many copies. Cells use these molecules to transform matter and energy, to respond to their environments, and to reproduce. As we mentioned in the opening story, these biological processes would not be possible outside the enclosure of a cell.
The cell theory, developed in the nineteenth century, recognizes this basic fact about life. It was the first unifying principle of biology and has three critical components:
- Cells are the fundamental units of life.
- All living organisms are composed of cells.
- All cells come from preexisting cells.
Cell theory has two important conceptual implications:
- Studying cell biology is in some sense the same as studying life. The principles that underlie the functions of a single bacterial cell are similar to those governing the approximately 60 trillion cells in an adult human.
- Life is continuous. All those human cells came from a single cell, a zygote (or fertilized egg). The zygote was formed when two cells fused: a sperm from the father and an egg from the mother. The cells of the parents’ bodies were all derived from their parents, and so on back through the generations—all the way back to the evolution of the first living cells.
Cell size can be limited by the surface area-to-volume ratio
Most cells are tiny. Their diameters range from about 1 to 100 micrometers (FIGURE 4.1). There are some exceptions: the eggs of birds are single cells that are, relatively speaking, enormous, and individual cells of several types of algae and bacteria are large enough to be viewed with the unaided eye.
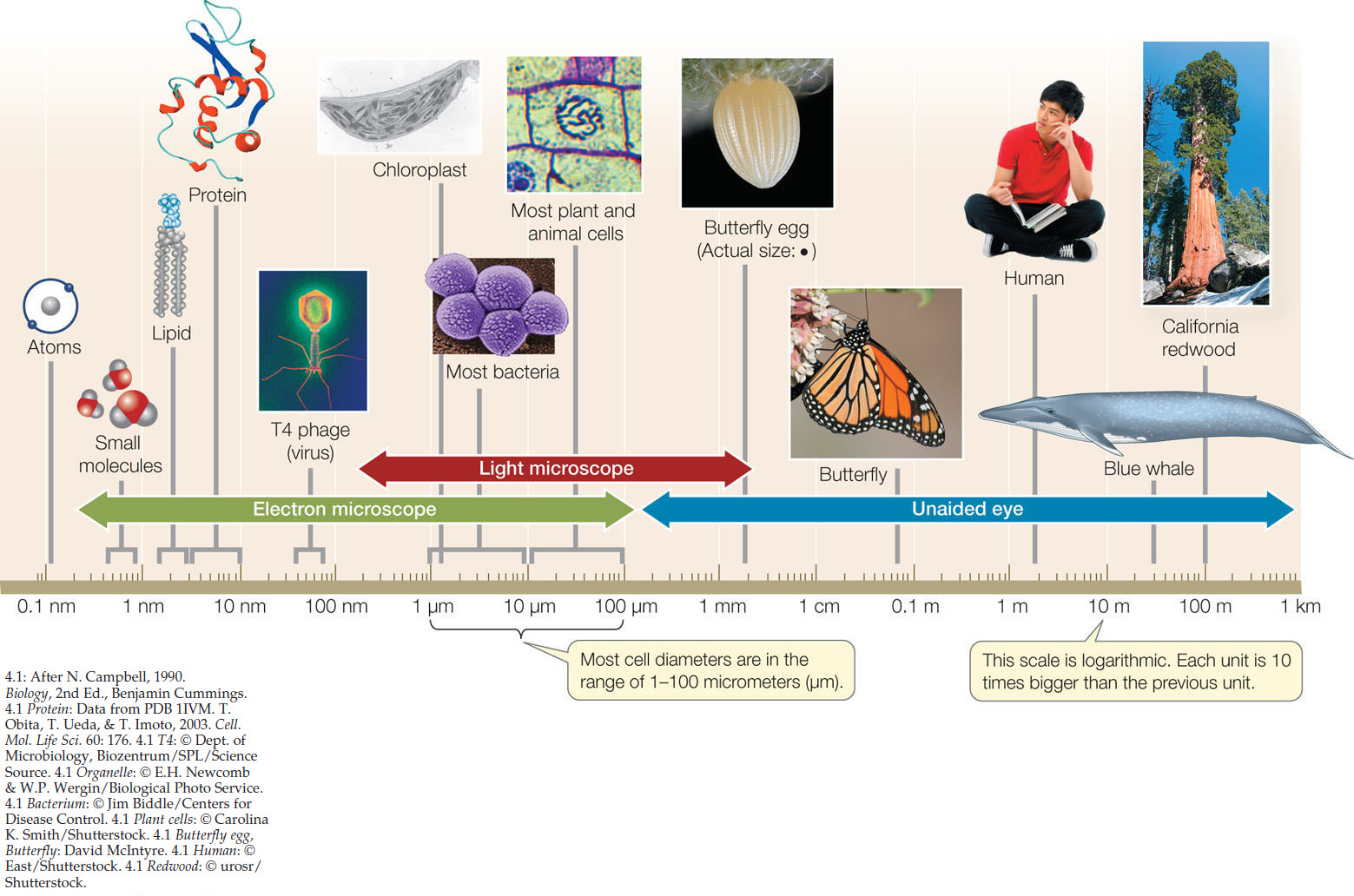
Small cell size is a practical necessity arising from the decrease in the surface area-to-volume ratio of any object as it increases in size. As an object increases in volume, its surface area also increases, but not as quickly (FIGURE 4.2). This phenomenon has biological significance for two reasons:
- The volume of a cell determines the amount of metabolic activity it carries out per unit of time.
- The surface area of a cell determines the amount of substances that can enter it from the outside environment, and the amount of waste products that can exit to the environment.
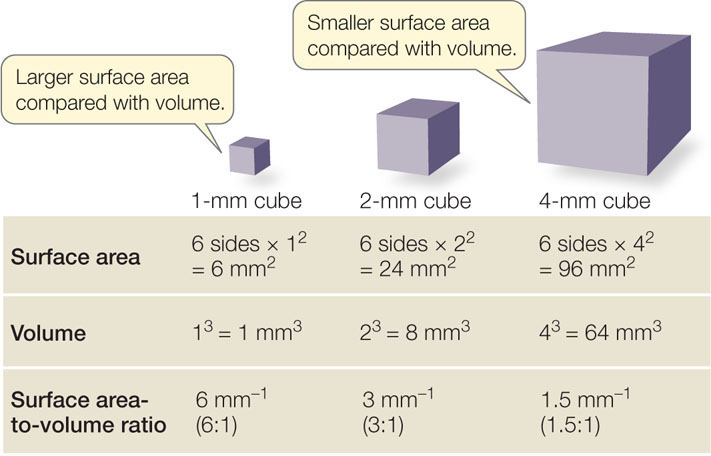
As a living cell grows larger, its metabolic activity, and thus its need for resources and its rate of waste production, increases faster than its surface area. In addition, substances must move from one location to another within the cell; the smaller the cell, the more easily this is accomplished. The large surface area-to-volume ratio represented by the many small cells of a multicellular organism enables it to carry out the many different functions required for survival.
62
However, for some cell types this general argument does not hold. Many cells are not shaped like cubes, but rather have an irregular shape; as they get larger, they can still have adequate exchange of materials with the environment by increasing their surface area by folds of their cell membrane. In other cases, cells can be quite large without greatly increased membrane surface. For instance, in a giraffe, nerve cells can be several meters long. In these cases, the rate of exchange of materials across the cell membrane must be increased, since the surface area is not adequately larger.
Cells can be studied structurally and chemically
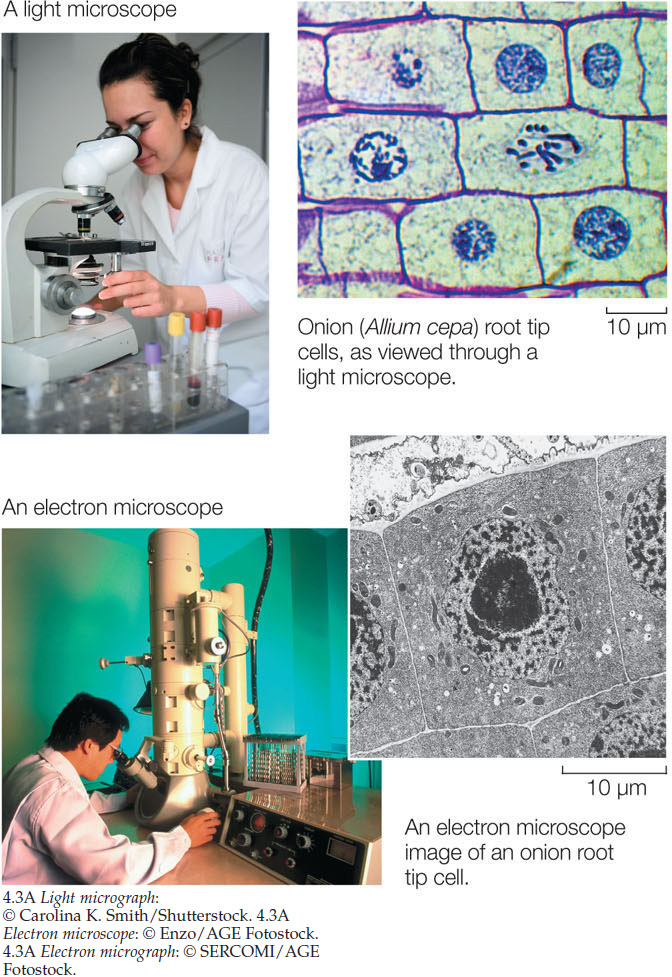
The small sizes of most cells necessitate special instruments to study them and their constituents. For visualizing cells, there are two types of microscopes (FIGURE 4.3):
- A light microscope uses glass lenses and visible light to form images. Structures that absorb more light are seen as darker than regions that do not, because the absorbed light does not reach the eye. The smallest detail that can be seen with such a microscope is about 0.2 μm in diameter, which is about 1,000 times smaller than an object the human eye can see. Light microscopes are used to visualize living cells and general cell structure.
- An electron microscope uses an electron beam focused by magnets to illuminate a specimen and produce an image on a TV-like screen. Structures that absorb the electrons appear darker than regions that do not. The size limit is 0.1 nm, which is 2 million times smaller than something the human eye can see. For electron microscopy, specimens must be preserved and stained using toxic heavy metals, so living cells cannot be visualized this way.
RESEARCH TOOLS
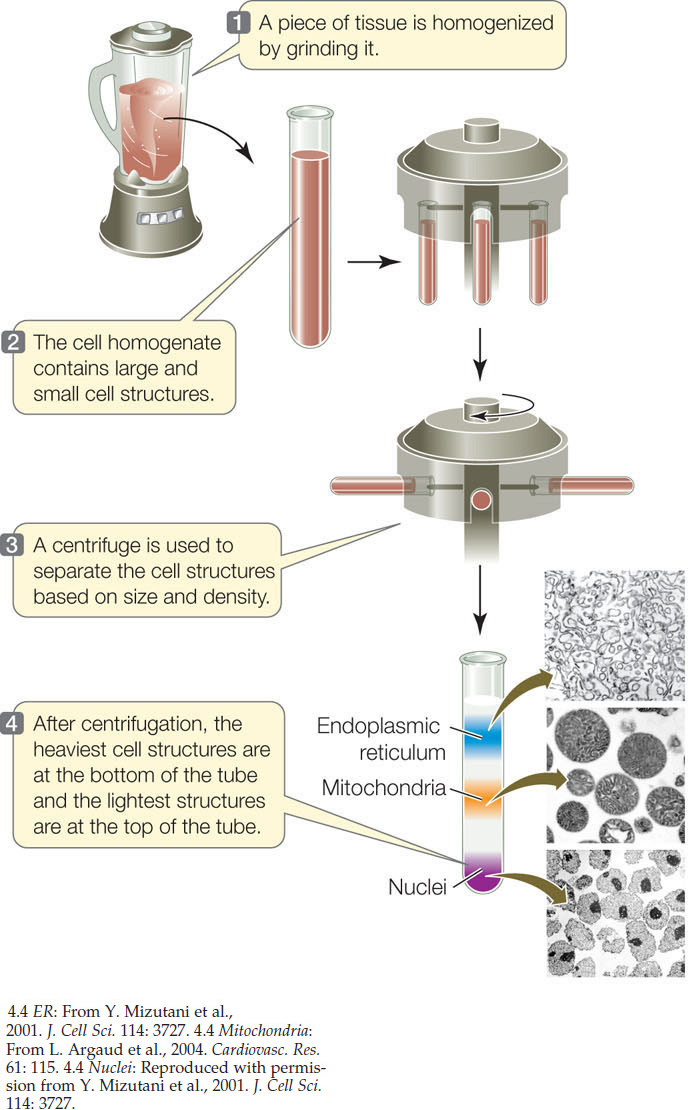
The chemical analysis of cells usually begins with breaking them open to make a cell-free extract. This can be done physically, using a blender or other homogenizing machinery, or by placing the cell in a chemical environment where it swells and bursts (see Figure 5.3). In either case, the resulting extract can be analyzed in terms of its composition and chemical reactions. For example, specific enzyme activities may be measured. If conditions are right in this test tube system, the properties of the cell-free extract are the same as those inside the cell. This last statement is of great importance, because it allows biologists to study the chemical processes that occur inside cells in the test tube, so that chemical changes can be easily measured.
A cell’s internal structures and even some of its macromolecules can be separated according to their sizes in a centrifuge that spins the tubes at a high speed (FIGURE 4.4). Once the subcellular structures are separated from one another, they are much easier to study.
RESEARCH TOOLS
The cell membrane forms the outer surface of every cell
As we will discuss at the end of this chapter, a key to the origin of cells was the enclosure of biochemical functions within a membrane. We will describe the cell membrane in more detail in Chapter 5, but for now, note that it consists of a phospholipid bilayer with proteins (see Figure 2.13B). Unless stained for light microscopy, the very thin (7 nm) cell membrane is visible only with electron microscopy. It has several important roles:
- The cell membrane acts as a selectively permeable barrier. preventing some substances from crossing it while permitting other substances to enter and leave the cell. In doing so, it allows the cell to maintain a stable internal environment that is distinct from the surrounding environment. This explains why a red blood cell contains the pigmented molecule hemoglobin but the surrounding blood plasma does not.
- As the cell’s boundary with the outside environment, the cell membrane is important in communicating with adjacent cells and receiving signals from the environment.
- The cell membrane often has proteins protruding from it that are responsible for binding and adhering to adjacent cells or to a surface. Thus the cell membrane plays an important structural role and contributes to cell shape.
Cells are classified as either prokaryotic or eukaryotic
Biologists classify all living things into three domains: Archaea, Bacteria, and Eukarya. The organisms in Archaea and Bacteria are collectively called prokaryotes because they have in common a prokaryotic cellular organization. A prokaryotic cell typically does not have membrane-enclosed internal compartments; in particular, it does not have a nucleus.
Eukaryotic cell organization is found in members of the domain Eukarya—the eukaryotes—which includes the protists (a diverse group of microorganisms), plants, fungi, and animals. In contrast to the prokaryotes, eukaryotes contain membrane-enclosed compartments called organelles where specific metabolic functions occur. The most notable of these is the cell nucleus, where most of the cell’s DNA is located and where gene expression begins:
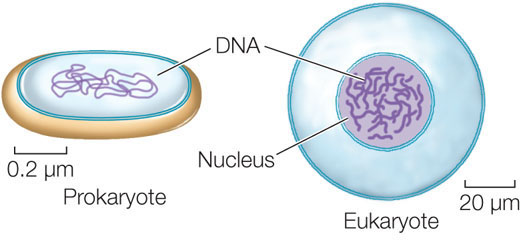
Just as a cell is an enclosed compartment that separates its contents from the surrounding environment, each organelle provides a compartment that separates certain molecules and biochemical reactions from the rest of the cell. This impressive “division of labor” provides possibilities for regulation and efficiency that were important in the evolution of complex organisms.
LINK
Eukaryotes arose from prokaryotes by endosymbiosis; see Concept 20.1
CHECKpointCONCEPT4.1
- In considering the origin of cells, why do biologists focus on the origin of the cell membrane?
- If a cell has a cube shape that is 500 μm on a side, what is its surface area-to-volume ratio? If the surface area-to-volume ratio should be more than 0.1 μm−1 for optimal cell function, would dividing this cell into 1 million individual cells (also cubes) meet this standard?
- What is the surface area-to-volume ratio of a giraffe’s nerve cell? [For simplicity, assume that it is tubular (a cylinder), with a length of 3 m and a diameter of 5 μm.]
- What evolutionary advantages does a eukaryotic cell have compared with a prokaryotic cell?
This section has introduced two structural themes in cell architecture: prokaryotic and eukaryotic. We’ll turn now to the organization of prokaryotic cells.