CONCEPT8.3 Genes Are Carried on Chromosomes
Genes are parts of chromosomes. More specifically, a gene is a sequence of DNA that resides at a particular site on a chromosome called a locus (plural loci). You have seen how the behavior of chromosomes during meiosis can explain Mendel’s laws of segregation (see Figure 8.3) and independent assortment (see Figure 8.6). However, the genetic linkage of genes on a single chromosome alters their pattern of inheritance.
Genetic linkage was first discovered in the fruit fly Drosophila melanogaster. This animal is an attractive experimental subject because it is small, easily bred, and has a short generation time (from fertilized egg to reproducing adult). In fact, the fruit fly has been a model organism for experimental genetics for more than a century. In Concept 8.1 we saw how Mendel successfully applied the scientific method to arrive at his laws of inheritance. Now we will examine the work of Thomas Hunt Morgan, who worked at Columbia University early in the twentieth century and used a similar approach to discover genetic linkage.
Genes on the same chromosome are linked, but can be separated by crossing over in meiosis
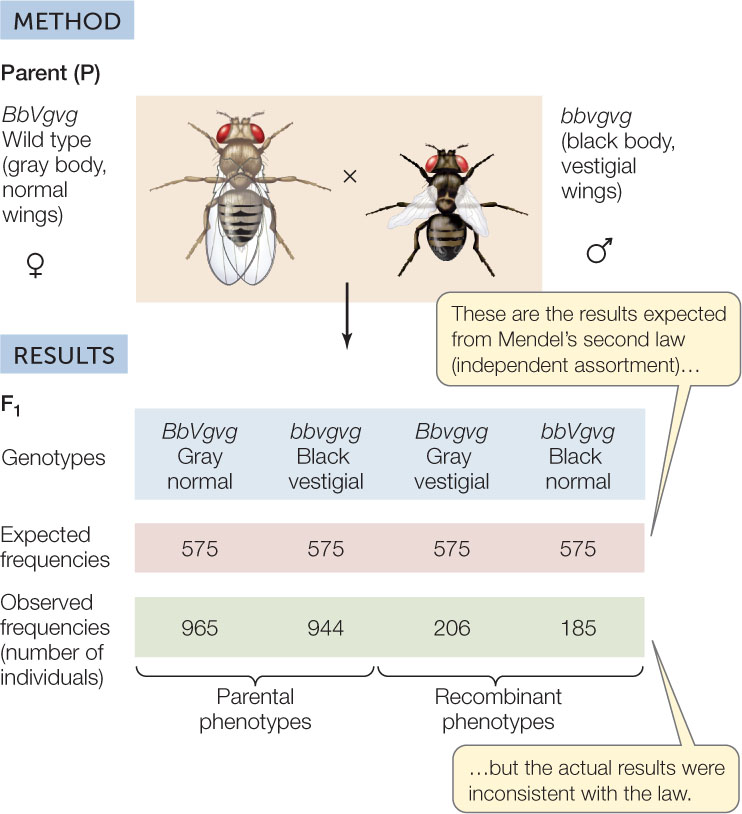
Some of the crosses Morgan performed with fruit flies yielded phenotypic ratios that were not in accordance with those predicted by Mendel’s law of independent assortment. Morgan did a test cross between flies with two known genotypes: BbVgvg and bbvgvg. The B and Vg genes control two characters, body color and wing shape:
- B (wild-type gray body) is dominant over b (black body).
- Vg (wild-type wing) is dominant over vg (vestigial, or very small, wing).
163
Morgan expected to see four phenotypes in a ratio of 1:1:1:1, but that is not what he observed. The body color gene and the wing size gene did not assort independently; instead, they were frequently inherited together, and most of the progeny showed one or the other of the parental phenotypes (FIGURE 8.13).
Investigation
HYPOTHESIS
Alleles for different characteristics always assort independently.
CONCLUSION
The hypothesis is rejected. These two genes do not assort independently, but are linked (on the same chromosome).
aT. H. Morgan. 1912. Science 36: 718–720
These results became understandable when Morgan considered the possibility that the two loci were linked on the same chromosome. Such genes would not be able to assort independently as predicted by Mendel’s second law. In this case, the test cross offspring might be expected to have only the parental phenotypes (gray flies with normal wings or black flies with vestigial wings) in a 1:1 ratio. If linkage were absolute, we would see only these two types of progeny. However, this did not happen. Why did some of Morgan’s flies show phenotypes different from their parents?
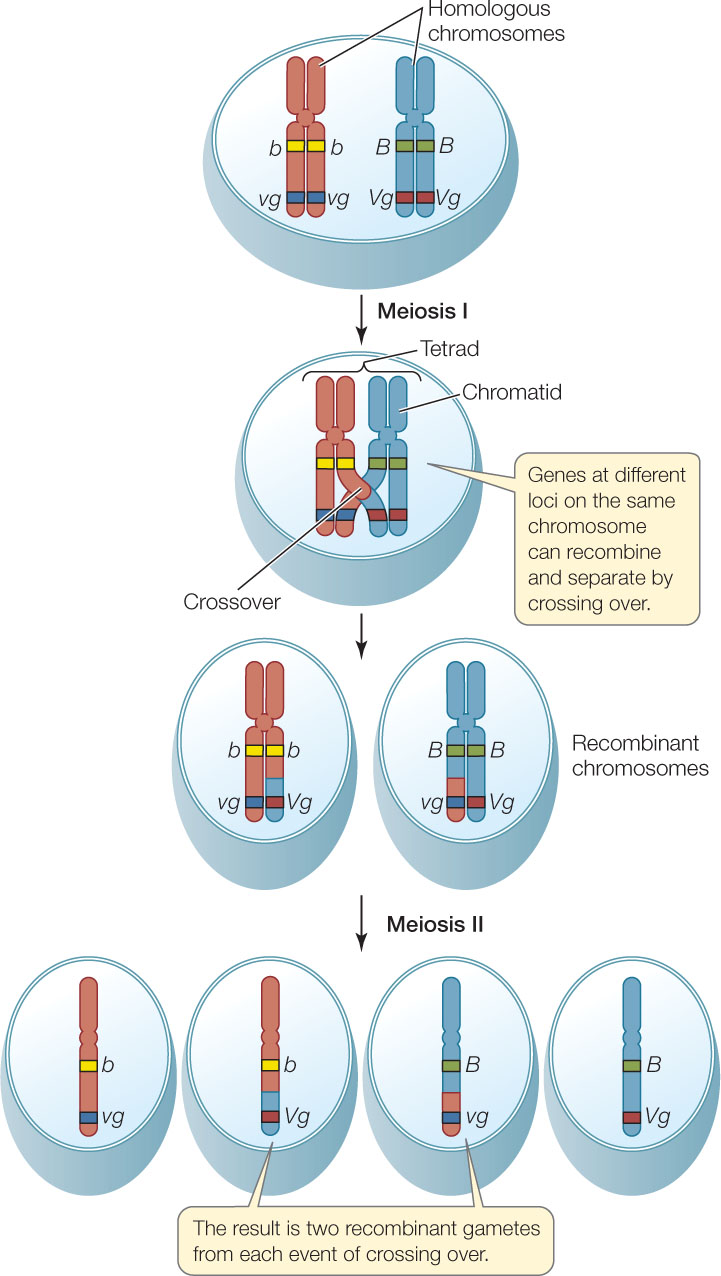
Some of Morgan’s flies displayed recombinant phenotypes because two homologous chromosomes can physically exchange corresponding segments during prophase I of meiosis—that is, by crossing over (FIGURE 8.14; see also Figure 7.13). Each exchange event involves two of the four chromatids in a tetrad—one from each member of the homologous pair—and can occur at any point along the length of the chromosome. The chromosome segments are exchanged reciprocally, so both chromatids become recombinant (that is, each chromatid ends up with genes from both of the organism’s parents).
164
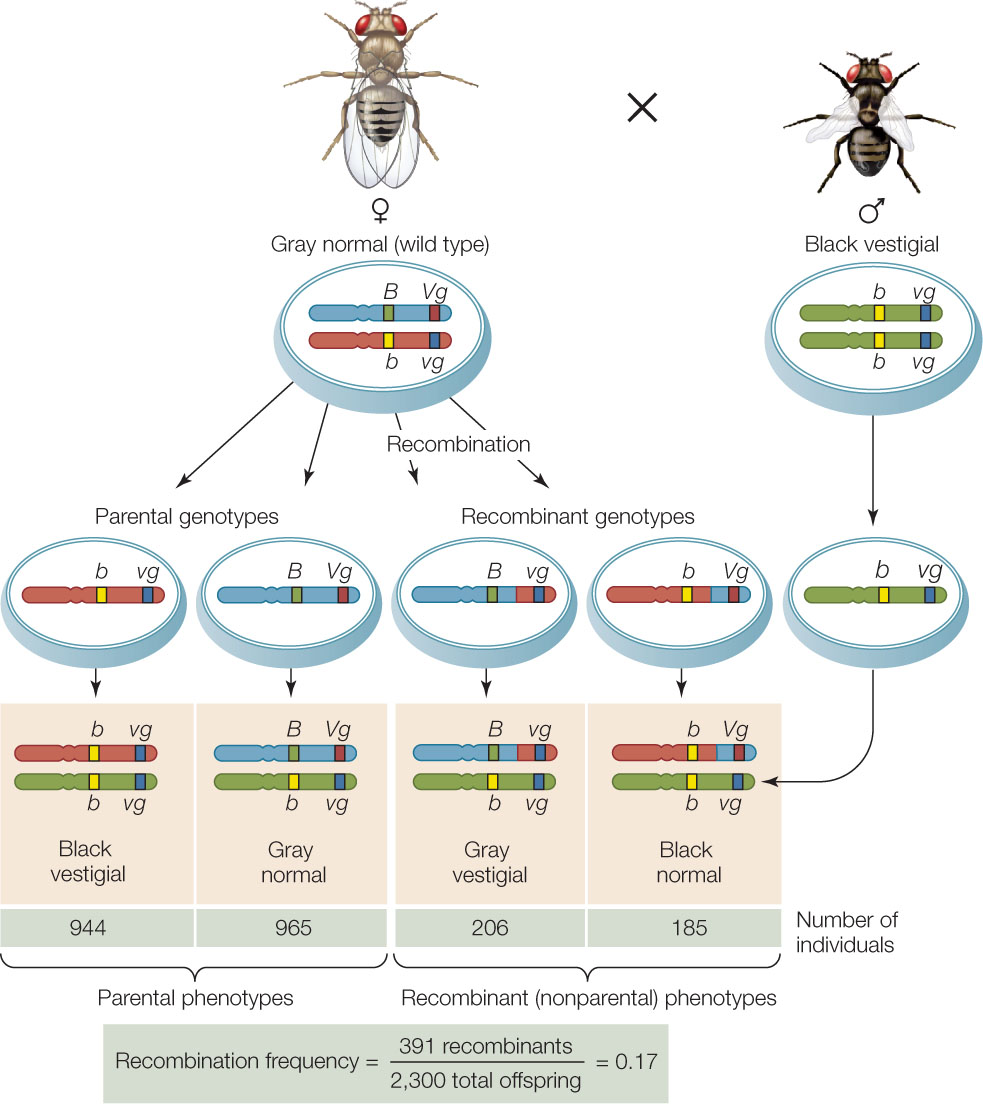
As a result of a crossing over event between two linked genes, not all the progeny of a cross have the parental phenotypes. Recombinant offspring appear as well, generally in proportions related to the recombination frequency between the two genes, which is calculated by dividing the number of recombinant progeny by the total number of progeny (FIGURE 8.15). Recombination frequencies are greater for loci that are farther apart on the chromosome than for loci that are closer together because crossing over is more likely to occur between genes that are far apart. By calculating recombination frequencies, geneticists can infer the locations of genes along a chromosome and generate a genetic map. Below is a map showing five genes on a fruit fly chromosome. It was constructed using the recombination frequencies generated by test crosses involving various pairs of the genes:
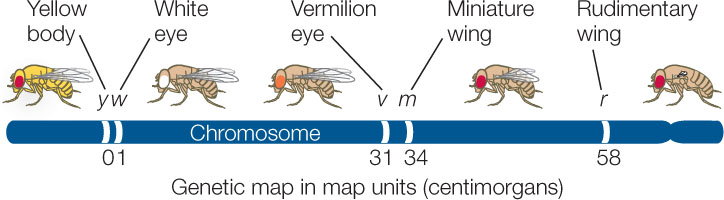
The recombination frequency between y and w is low, so they are close together on the map. Recombination between y and v is more frequent, so they are farther apart. The recombination frequencies are converted to map units (also called centimorgans, cM); one map unit is equivalent to an average recombination frequency of 0.01 (one percent).
The era of gene sequencing has made mapping less important in some areas of genetics research. However, mapping is still one way to verify that a particular DNA sequence corresponds with a particular phenotype. The phenomenon of linkage has allowed biologists to isolate genes and to create genetic markers that are linked to important genes, making it easy to identify individuals carrying particular alleles. This is particularly important in breeding new crops and animals for agriculture, and for identifying humans carrying medically significant mutations.
Linkage is also revealed by studies of the X and Y chromosomes
The fruit fly genome has four pairs of chromosomes: in three pairs, the chromosomes are similar in size to one another and are called autosomes. The fourth pair has two chromosomes of different sizes. These determine the sex of the fly and are called the sex chromosomes, shown here in green:
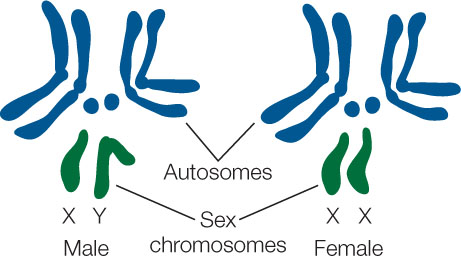
Note that the female fly has two X chromosomes and that the male has only one, the other being the Y chromosome: females are XX and males are XY. It turns out that in addition to being different sizes, many genes on the X chromosome are not present on the Y. This means that males have only one copy of these genes. The X chromosome was one of the first to have specific genes assigned to it.
Morgan identified a gene that controls eye color in Drosophila. The wild-type allele of the gene confers red eyes, whereas a recessive mutant allele confers white eyes. Morgan’s experimental crosses with flies carrying the mutant allele demonstrated that this eye color locus is on the X chromosome. If we abbreviate the eye color alleles as R (red eyes) and r (white eyes), the presence of the alleles on the X chromosome is designated by XR and Xr.
165
Morgan crossed a homozygous red-eyed female (XRXR) with a white-eyed male. The male is designated XrY because the Y does not carry any allele for this gene. (Any gene that is present as a single copy in a diploid organism is called hemizygous. A male will express all the alleles of his one X chromosome, whether or not they are dominant.) All the sons and daughters from this cross had red eyes, because red (R) is dominant over white (r) and all the progeny had inherited a wild-type X chromosome (XR) from their mother (FIGURE 8.16A). Note that this phenotypic outcome would have occurred even if the R gene had been present on an autosome rather than a sex chromosome. In that case, the male would have been homozygous recessive—rr.
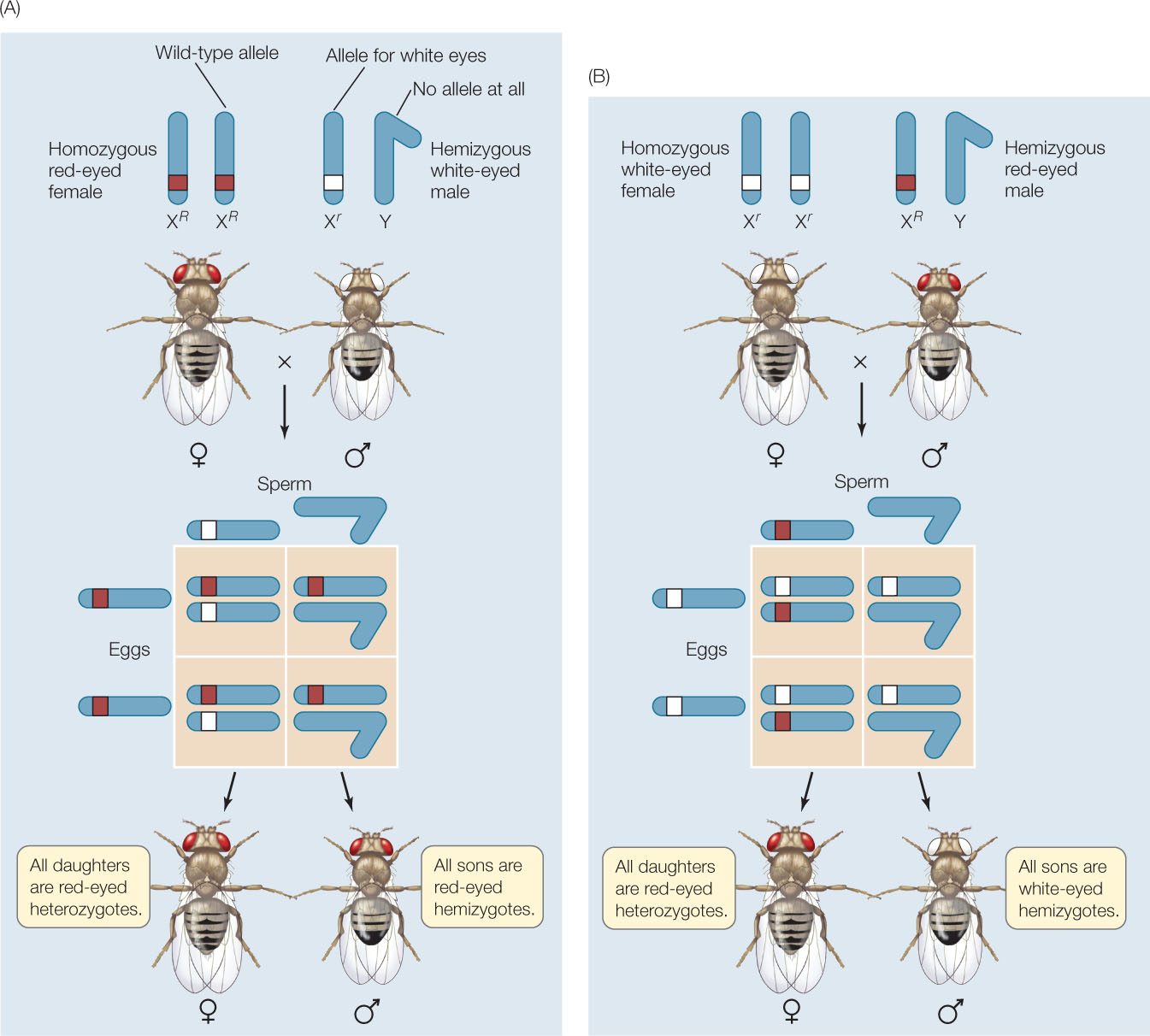
When Morgan performed the reciprocal cross, in which a white-eyed female (XrXr) was mated with a red-eyed male (XRY), the results were unexpected: all the sons were white-eyed and all the daughters were red-eyed (FIGURE 8.16B). The sons from the reciprocal cross inherited their only X chromosome from their white-eyed mother and were therefore hemizygous for the white allele. The daughters, however, got an X chromosome bearing the white allele from their mother and an X chromosome bearing the red allele from their father; therefore they were red-eyed heterozygotes. When these heterozygous females were mated with red-eyed males, half their sons had white eyes but all their daughters had red eyes. Together, these results showed that eye color was carried on the X chromosome and not on the Y.
These and other experiments led to the term sex-linked inheritance: inheritance of a gene that is carried on a sex chromosome. (This term is misleading because “sex-linked” inheritance is not really linked to the sex of an organism—after all, both males and females carry X chromosomes.) In mammals, the X chromosome is larger and carries more genes than the Y. For this reason, most examples of sex-linked inheritance involve genes that are carried on the X chromosome.
166
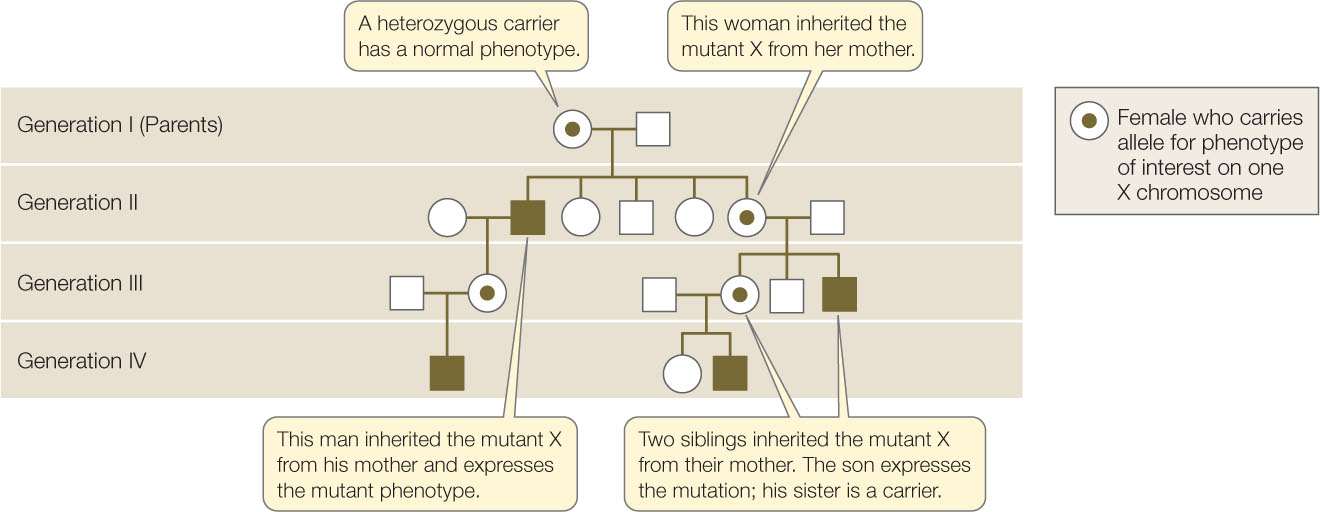
Many sexually reproducing species, including humans, have sex chromosomes. As in fruit flies, human males are XY, females are XX, and relatively few of the genes that are present on the X chromosome are present on the Y. Pedigree analyses of X-linked recessive phenotypes like the one in FIGURE 8.17 reveal the following patterns (compare with the pedigrees of non-X-linked phenotypes in Figure 8.8):
- The phenotype appears much more often in males than in females, because only one copy of the allele is needed for its expression in males, whereas two copies must be present in females.
- A male with the mutation can pass it on only to his daughters; all his sons get his Y chromosome.
- Daughters who receive one X-linked mutation are heterozygous carriers. They are phenotypically normal, but they can pass the mutant allele to their sons or daughters. On average, half their children will inherit the mutant allele since half of their X chromosomes carry the normal allele.
- The mutant phenotype can skip a generation if the mutation passes from a male to his daughter (who will be phenotypically normal) and then to her son.
Some genes are carried on chromosomes in organelles
The nucleus is not the only organelle in a eukaryotic cell that carries genetic material. Mitochondria and plastids (including chloroplasts) each contain several copies of a small chromosome that carries a small number of genes. For example, in humans there are about 21,000 genes in the nuclear genome and 37 in the mitochondrial genome. Plastids have five times as many genes as mitochondria. But note that these organelle genomes do not encode all of the molecules that make up the organelle. Most of the proteins and some of the RNAs in organelles are encoded by the nuclear genome and imported from the cytoplasm.
Genes are carried on chromosomes
The pedigree shows the inheritance pattern of a rare mutant phenotype in humans, congenital cataract (filled-in symbols).
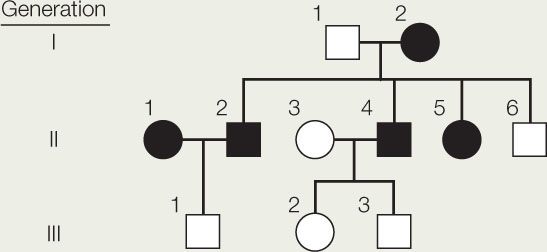
- Are cataracts inherited as autosomal dominant? Autosomal recessive? Sex-linked dominant? Sex-linked recessive?
- Person #5 in the second generation marries a man who does not have cataracts. Two of their four children, a boy and a girl, develop cataracts. What is the probability that their next child will be a girl with cataracts?
The inheritance of organelle genes differs from that of nuclear genes because in most organisms, mitochondria and plastids are inherited only from the mother. Most egg cells in plants and animals contain abundant cytoplasm and organelles, but the only part of the sperm that survives to take part in the union of haploid gametes is the nucleus. You have inherited your mother’s mitochondria (with their genes) but not your father’s. The inheritance of organelles and their genes is therefore non-Mendelian and is described as maternal or cytoplasmic inheritance, since the inherited organelles come from the maternal cytoplasm.
167
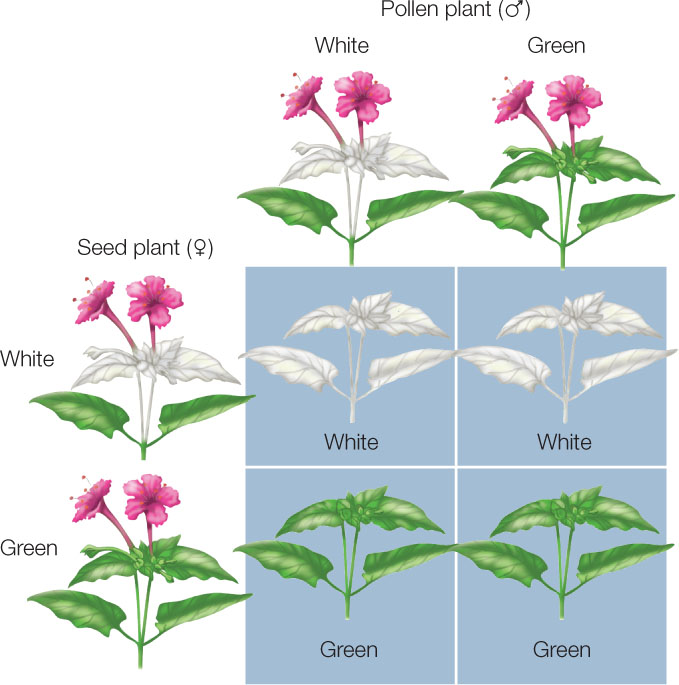
Some of the genes carried by cytoplasmic organelles are important for organelle assembly and function, and mutations of these genes can have profound effects on the organism. For example, in plants, certain plastid gene mutations affect the proteins that assemble chlorophyll molecules into photosystems (see Figure 6.19). These mutations result in a phenotype that is essentially white instead of green. FIGURE 8.18 illustrates the cytoplasmic inheritance of a mutant plastid gene that confers white color to leaves.
CHECKpointCONCEPT8.3
- Describe the differences in patterns of inheritance between a gene present in the nucleus and a gene present in the mitochondria.
- Explain the concept of linkage. If you performed a test cross with a fruit fly that is heterozygous for two genes, how would you conclude that the two genes are linked?
- Red-green color blindness is inherited as a sex-linked recessive. Two parents with normal color vision have a child who is red-green color-blind. Is the child a boy or girl? Draw a pedigree of the family. Draw a Punnett square to show gametes and offspring with regard to the X and Y chromosomes and the normal and color-blind alleles.
Like eukaryotes, prokaryotes contain genes that determine their phenotypes. Sexual reproduction in eukaryotes involves two sets of chromosomes and meiosis, giving rise to haploid gametes. Let’s look next at how reproduction and inheritance differ in prokaryotes, which are haploid.