CONCEPT17.3 Speciation May Occur through Geographic Isolation or in Sympatry
Many scientists who study speciation have concentrated on geographic processes that result in the splitting of an ancestral species. Splitting the range of a species is one obvious way of achieving such a division, but it is not the only way.
Physical barriers give rise to allopatric speciation
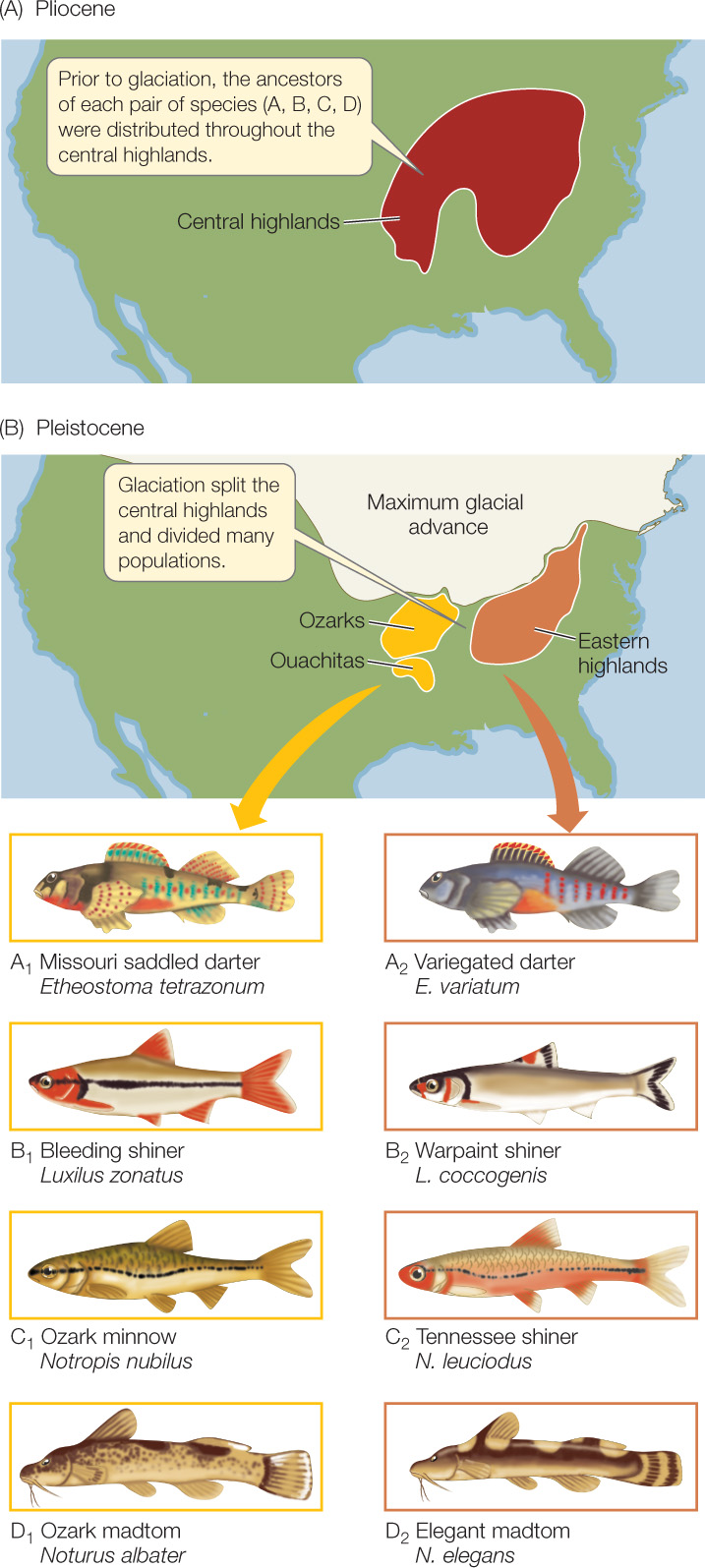
Speciation that results when a population is divided by a physical barrier is known as allopatric speciation (Greek allos, “other”; patria, “homeland”). Allopatric speciation is thought to be the dominant mode of speciation in most groups of organisms. The physical barrier that divides the range of a species may be a body of water or a mountain range for terrestrial organisms, or dry land for aquatic organisms—in other words, any type of habitat that is inhospitable to the species. Such barriers can form when continents drift, sea levels rise and fall, glaciers advance and retreat, or climates change. The populations separated by such barriers are often, but not always, initially large. The lineages that descend from these founding populations evolve differences for a variety of reasons, including mutation, genetic drift, and adaptation to different environments in the two areas. As a result, many pairs of closely related sister species—species that are each other’s closest relatives—may exist on opposite sides of the geographic barrier. An example of a physical geographical barrier that produced many pairs of sister species was the Pleistocene glaciation that isolated freshwater streams in the eastern highlands of the Appalachian Mountains from streams in the Ozark and Ouachita mountains about 20,000–80,000 years ago (FIGURE 17.6). This splitting event resulted in many parallel speciation events among isolated lineages of stream-dwelling organisms.
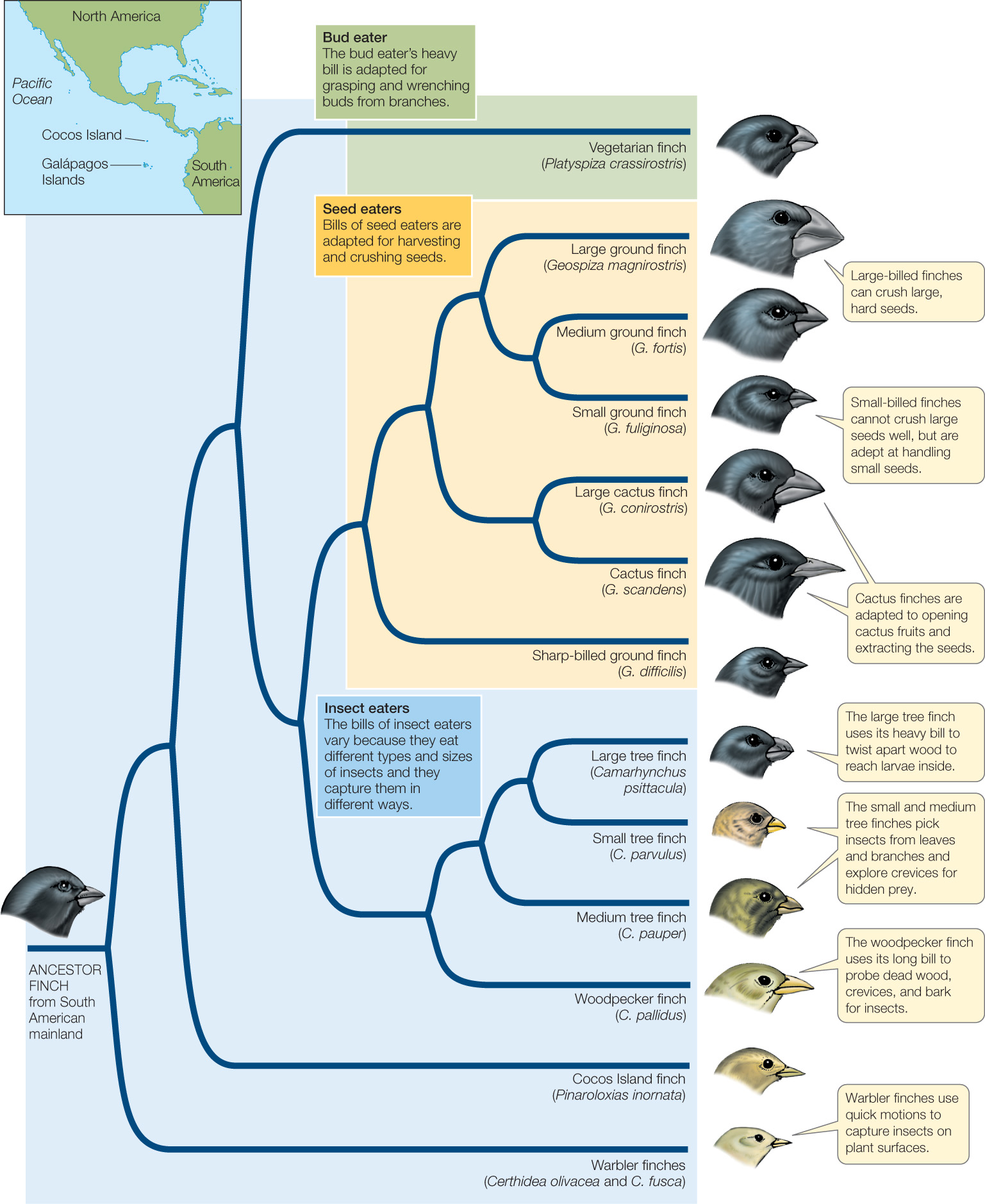
Allopatric speciation also may result when some members of a population cross an existing barrier and establish a new, isolated population. The 14 species of finches found in the Galápagos archipelago some 1,000 kilometers off the coast of Ecuador are the result of this process. Darwin’s finches (as they are usually called, because Darwin was the first scientist to study them) arose in the Galápagos from a single South American finch species that colonized the islands. Today the Galápagos species differ strikingly from their closest mainland relative and from one another (FIGURE 17.7). The islands of the archipelago are sufficiently far apart that the finches move among them only infrequently. In addition, environmental conditions differ widely from island to island. Some islands are relatively flat and arid; others have forested mountain slopes. Over millions of years, finch lineages on the different islands have differentiated to the point that when occasional immigrants arrive from other islands, they either do not breed with the residents, or if they do, the resulting offspring do not survive as well as the offspring of established residents. The genetic distinctness of each finch species from the others and the genetic cohesiveness of the individual species are thus maintained.
349
350
Speciation may occur through geographic isolation
The different species of Darwin’s finches shown in the phylogeny in Figure 17.7 have all evolved on islands of the Galápagos archipelago within the past 3 million years. Molecular clock analysis (see Concept 16.3) has been used to determine the dates of the various speciation events in that phylogeny. Geological techniques for dating rock samples (see Concept 18.1) have been used to determine the ages of the various Galápagos islands. The table shows the number of species of Darwin’s finches and the number of islands that have existed in the archipelago at several times during the past 4 million years.
- Plot the number of species of Darwin’s finches and the number of islands in the Galápagos archipelago (dependent variables) against time (independent variable).
- Are the data consistent with the hypothesis that isolation of populations on newly formed islands is related to speciation in this group of birds? Why or why not?
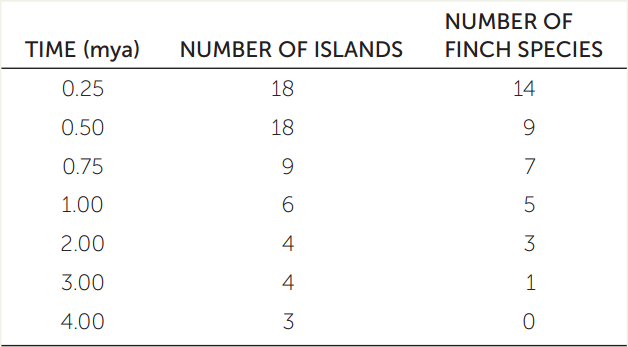
- If no more islands form in the Galápagos archipelago, do you think that speciation by geographic isolation will continue to occur among Darwin’s finches? Why or why not? What additional data could you collect to test your hypothesis (without waiting to see if speciation occurs)?
Sympatric speciation occurs without physical barriers
Geographic isolation is usually required for speciation, but under some circumstances speciation can occur in the absence of a physical barrier. Speciation without physical isolation is called sympatric speciation (Greek sym, “together with”). But how can such speciation happen? Given that speciation is usually a gradual process, how can reproductive isolation develop when individuals have frequent opportunities to mate with one another?
Sympatric speciation may occur with some forms of disruptive selection (see Concept 15.4) in which individuals with certain genotypes have a preference for distinct microhabitats where mating takes place. The opening story of African cichlids was an example. Another example appears to be taking place in the apple maggot fly (Rhagoletis pomonella) of eastern North America. Until the mid-1800s, Rhagoletis flies courted, mated, and deposited their eggs only on hawthorn fruits. About 150 years ago, some flies began to lay their eggs on apples, which European immigrants had introduced into eastern North America. Apple trees are closely related to hawthorns, but the smell of the fruits differs, and the apple fruits appear earlier than those of hawthorns. Some early-emerging female Rhagoletis laid their eggs on apples, and over time, a genetic preference for the smell of apples evolved among early-emerging flies. When the offspring of these flies sought out apple trees for mating and egg deposition, they mated with other flies reared on apples, which shared the same preferences.
Today the two groups of Rhagoletis pomonella in the eastern U.S. appear to be on the way to becoming distinct species. One group mates and lays eggs primarily on hawthorn fruits, the other on apples. The incipient species are partially reproductively isolated because they mate primarily with individuals raised on the same fruit and because they emerge from their pupae at different times of the year. In addition, the apple-feeding flies now grow more rapidly on apples than they originally did. Sympatric speciation that arises from such host-plant specificity may be widespread among insects, many of which feed only on a single plant species.
The most common means of sympatric speciation, however, is polyploidy—the duplication of sets of chromosomes within individuals. Polyploidy can arise either from chromosome duplication in a single species (autopolyploidy) or from the combining of the chromosomes of two different species (allopolyploidy).
An autopolyploid individual originates when, for example, two accidentally unreduced diploid gametes (with two sets of chromosomes) combine to form a tetraploid individual (with four sets of chromosomes). Tetraploid and diploid individuals of the same species are reproductively isolated because their hybrid offspring are triploid. Even if these offspring survive, they are usually sterile; they cannot produce normal gametes because their chromosomes do not segregate evenly during meiosis. So a tetraploid individual cannot produce fertile offspring by mating with a diploid individual—but it can do so if it self-fertilizes or mates with another tetraploid. Thus polyploidy can result in complete reproductive isolation in two generations—an important exception to the general rule that speciation is a gradual process.
Allopolyploids may be produced when individuals of two different (but closely related) species interbreed. Such hybridization often disrupts normal meiosis, which can result in chromosomal doubling. Allopolyploids are often fertile because each of the chromosomes has a nearly identical partner with which to pair during meiosis.
Speciation by polyploidy has been particularly important in the evolution of plants, although it has contributed to speciation in animals as well (such as the example in Figure 17.2). Botanists estimate that about 70 percent of flowering plant species and 95 percent of fern species are the result of recent polyploidization. Some of these species arose from hybridization between two species followed by chromosomal duplication and self-fertilization. Other species diverged from polyploid ancestors, so that the new species shared their ancestors’ duplicated sets of chromosomes. New species arise by polyploidy more easily among plants than among animals because plants of many species can reproduce by self-fertilization. In addition, if polyploidy arises in several offspring of a single parent, the siblings can fertilize one another.
351
CHECKpointCONCEPT17.3
- Explain how speciation via polyploidy can happen in only two generations.
- What are some obstacles to sympatric speciation?
- If allopatric speciation is the most prevalent mode of speciation, what do you predict about the geographic distributions of many closely related species? Does your answer differ for species that are sedentary versus species that are highly mobile?
Most populations separated by a physical barrier become reproductively isolated only slowly and gradually. If two incipient species once again come into contact with each other, what keeps them from merging back into a single species?