Concept 19.3: Ecological Communities Depend on Prokaryotes
Prokaryotic cells and their associations do not usually live in isolation. Rather, they live in communities of many different species, often including microscopic eukaryotes. Whereas some microbial communities are harmful to humans, others provide important services. They help us digest our food, break down municipal waste, and recycle organic matter and chemical elements in the environment.
Many prokaryotes form complex communities
Some microbial communities form layers in sediments, and others form clumps a meter or more in diameter. Many microbial communities tend to form dense biofilms. Upon contacting a solid surface, the cells bind to that surface and secrete a sticky, gel-like polysaccharide matrix that traps other cells (FIGURE 19.19). Once a biofilm forms, the cells become more difficult to kill.
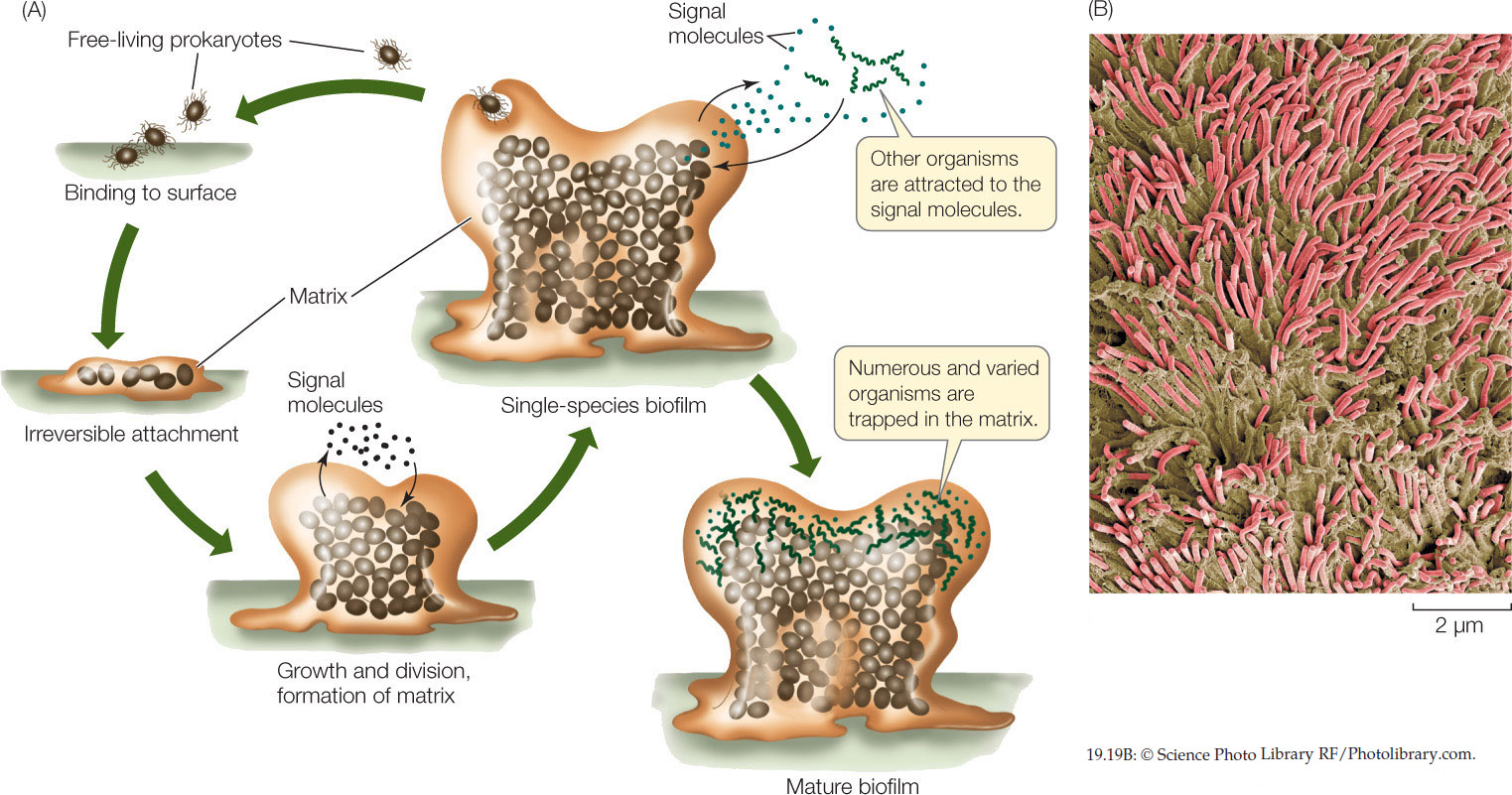
Biofilms are found in many places, and in some of those places they cause problems for humans. The material on our teeth that we call dental plaque is a biofilm. Pathogenic bacteria are difficult for the immune system—and modern medicine—to combat once they form a biofilm, which may be impermeable to antibiotics. Worse, some drugs stimulate the bacteria in a biofilm to lay down more matrix, making the film even more impermeable. Biofilms may form on just about any available surface, including contact lenses and artificial joint replacements. They foul metal pipes and cause corrosion, a major problem in steam-driven electricity generation plants. Fossil stromatolites—large, rocky structures made up of alternating layers of fossilized biofilm and calcium carbonate—are among the oldest remnants of life on Earth (see Figure 18.6B).
391
Some biologists are studying the chemical signals that prokaryotes use to communicate with one another and that trigger density-linked activities such as biofilm formation. We saw one example of this type of communication—called quorum sensing—in the chapter-opening discussion of bioluminescent Vibrio. In the case of health-threatening bacteria, researchers hope to find ways to block the quorum-sensing signals that lead to the production of the matrix polysaccharides, thus preventing pathological biofilms from forming.
Microbiomes are critical to the health of many eukaryotes
Although only a few bacterial species are pathogens, popular notions of bacteria as “germs” and fear of the consequences of infection cause many people to assume that most bacteria are harmful. Increasingly, however, biologists are discovering that the health of humans (as well as that of other eukaryotes) depends in large part on the health of our microbiomes: the communities of bacteria and archaea that inhabit our bodies.
Every surface of your body is covered with diverse communities of bacteria. The Human Microbiome Project has identified more than 10,000 species living in and on humans. Inside your body, your digestive system teems with bacteria (FIGURE 19.20). If you count up all the cells in a human body, only about 10 percent of them are human cells. The rest are microbes—mostly bacteria, along with some archaea and microscopic eukaryotes. When these communities are disrupted, they must be restored before the body can function normally.
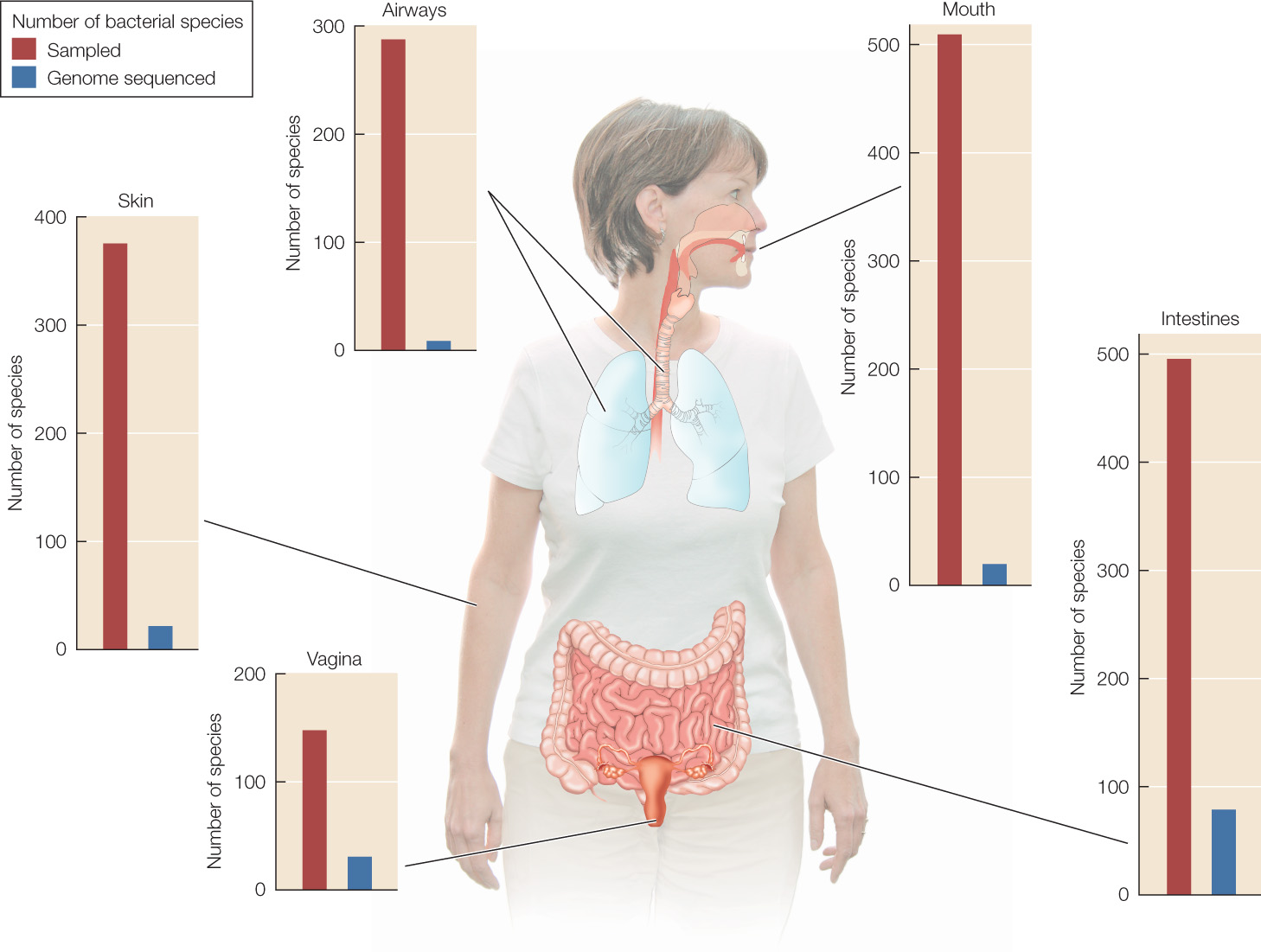
Biologists are discovering that many complex health problems are linked to the disruption of our microbiomes. These diverse microbial communities affect the expression of our genes and play a critical role in the development and maintenance of a healthy immune system. When our microbiomes contain an appropriate community of beneficial species, our bodies function normally. But these communities are strongly affected by our life experiences, by the food we eat, by the medicines we take, and by our exposure to various environmental toxins. The recent rapid increase in the rate of autoimmune diseases in humans—diseases in which the immune system begins to attack the body—has been linked to the changing diversity and composition of our microbiomes.
The early acquisition of an appropriate microbiome is critical for lifelong health. Normally, a human infant acquires much of its microbiome at birth, from the microbiome in its mother’s vagina. Other components of the microbiome are also acquired from the mother, especially through breast feeding. Recent studies have shown that babies born by cesarean section, as well as babies that are bottle-fed on artificial milk formula, typically acquire microbes from a wider variety of sources. Many of the bacteria acquired in this way are not well suited for human health. Biologists have discovered that the incidence of many autoimmune diseases is higher in people who were born by cesarean section and in those who were fed on formula as infants, compared with individuals who were born vaginally and breast-fed as infants. The difference appears to be related to the composition of the individual’s original microbiome.
392
Our microbiomes may be related to many other health concerns as well. For example, physicians have long noted a connection between autism and gastrointestinal disorders in humans. In 2012, microbiologists discovered that children with autism have high levels of a genus of bacteria known as Sutterella adherent to their intestinal walls. These bacteria are absent or rare in children without autism. It is not yet known if these bacteria are causing the gastrointestinal problems, or if they are merely symptoms of the gastrointestinal disorders, but there is growing evidence that intestinal microbiomes are distinct in children with autism.
Humans require some of the metabolic products—especially vitamins B12 and K—produced by the microbiome living in the large intestine. Communities of bacteria line our intestines with a dense biofilm that is in intimate contact with the mucosal lining of the gut. This biofilm facilitates nutrient transfer from the intestine into the body, functioning like a specialized “tissue” that is essential to our health. This biofilm has a complex ecology that scientists have just begun to explore in detail—including the possibility that the species composition of an individual’s gut microbiome may contribute to obesity (or the resistance to it).
Animals also harbor a variety of bacteria and archaea in their digestive tracts, many of which play important roles in digestion. Cattle depend on prokaryotes to break down plant material. Like most animals, cattle cannot produce cellulase, the enzyme needed to start the digestion of the cellulose that makes up the bulk of their plant food. However, bacteria living in a special section of the gut, called the rumen, produce enough cellulase to process the daily diet for the cattle (see Concept 30.2).
393
Marshall and Warren set out to satisfy Koch’s postulates:
Test 1
The microorganism must be present in every case of the disease.
Results: Biopsies from the stomachs of many patients revealed that the bacterium was always present if the stomach was inflamed or ulcerated.
Test 2
The microorganism must be cultured from a sick host.
Results: The bacterium was isolated from biopsy material and eventually grown in culture media in the laboratory.
Test 3
The isolated and cultured bacteria must be able to induce the disease.
Results: Marshall was examined and found to be free of bacteria and inflammation in his stomach. After drinking a pure culture of the bacterium, he developed stomach inflammation (gastritis).
Test 4
The bacteria must be recoverable from newly infected individuals.
Results: Biopsy of Marshall’s stomach 2 weeks after he ingested the bacteria revealed the presence of the bacterium, now christened Helicobacter pylori, in the inflamed tissue.
Conclusion
Antibiotic treatment eliminated the bacteria and the inflammation in Marshall. The experiment was repeated on healthy volunteers, and many patients with gastric ulcers were cured with antibiotics. Thus Marshall and Warren demonstrated that the stomach inflammation leading to ulcers is caused by H. pylori infections in the stomach.
A small minority of bacteria are pathogens
The late nineteenth century was a productive era in the history of medicine—a time when bacteriologists, chemists, and physicians proved that many diseases are caused by microbial agents. During this time, the German physician Robert Koch laid down a set of four rules for establishing that a particular microorganism causes a particular disease:
- The microorganism is always found in individuals with the disease.
- The microorganism can be taken from the host and grown in pure culture.
- A sample of the culture produces the same disease when injected into a new, healthy host.
- The newly infected host yields a new, pure culture of microorganisms identical to those obtained in the second step.
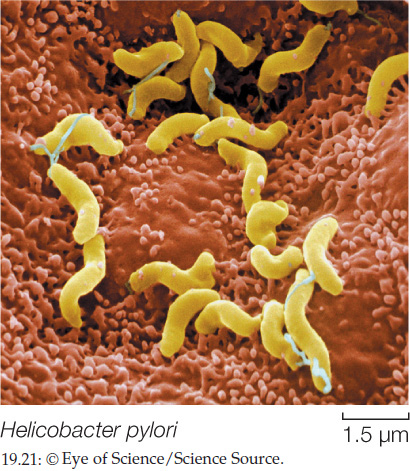
These rules, called Koch’s postulates, were important tools in a time when it was not widely understood that microorganisms cause disease. Although modern medical science has more powerful diagnostic tools, Koch’s postulates remain useful. For example, physicians were taken aback in the 1990s when stomach ulcers—long accepted and treated as the result of excess stomach acid—were shown by Koch’s postulates to be caused by the bacterium Helicobacter pylori (FIGURE 19.21).
For an organism to be a successful pathogen, it must:
- arrive at the body surface of a potential host;
- enter the host’s body;
- evade the host’s defenses;
- reproduce inside the host; and
- infect a new host.
Failure to complete any of these steps ends the reproductive career of a pathogenic organism. Yet in spite of the many defenses available to potential hosts (see Chapter 39), some bacteria are very successful pathogens. Pathogenic bacteria are often surprisingly difficult to combat, even with today’s arsenal of antibiotics. One source of this difficulty is their ability to form biofilms.
APPLY THE CONCEPT: A small minority of bacteria are pathogens
Imagine you are in charge of maintaining a trout hatchery. Some trout are exhibiting loss of tissue at the tips of their fins, and you suspect a bacterial infection. You isolate and culture two species of bacteria from a trout with affected fins.
- How would you satisfy the first of Koch’s postulates?
- Imagine that bacterium 1 satisfies the first of Koch’s postulates, but bacterium 2 does not. What presence or absence of data from the cultures from many samples of infected fish would lead you to this conclusion?
- You already know that bacterium 1 satisfies the second of Koch’s postulates. You decide to conduct a test of the third postulate. To your surprise, the test animals are all healthy and show no sign of disease. What are some possible explanations? How would you test your hypotheses?
394
For the host, the consequences of a bacterial infection depend on several factors. One is the invasiveness of the pathogen: its ability to multiply in the host’s body. Another is its toxigenicity: its ability to produce toxins (chemical substances that are harmful to the host’s tissues). Corynebacterium diphtheriae, the agent that causes diphtheria, has low invasiveness and multiplies only in the throat, but its toxigenicity is so great that the entire body is affected. In contrast, Bacillus anthracis, which causes anthrax, has low toxigenicity but is so invasive that the entire bloodstream ultimately teems with the bacteria.
There are two general types of bacterial toxins: endotoxins and exotoxins. Endotoxins are released when certain Gram-negative bacteria grow or lyse (burst). Endotoxins are lipopoly-saccharides (complexes consisting of a polysaccharide and a lipid component) that form part of the outer bacterial membrane. Endotoxins are rarely fatal to the host; they normally cause fever, vomiting, and diarrhea. Among the endotoxin producers are some strains of the proteobacteria Salmonella and Escherichia.
Exotoxins are soluble proteins released by living, multiplying bacteria. They are highly toxic—sometimes fatal—to the host. Human diseases induced by bacterial exotoxins include tetanus (Clostridium tetani), cholera (Vibrio cholerae), and bubonic plague (Yersinia pestis). Anthrax is caused by three exotoxins produced by Bacillus anthracis. Botulism is caused by exotoxins produced by Clostridium botulinum, which are among the most poisonous ever discovered. The lethal dose for humans of one exotoxin of C. botulinum is about one-millionth of a gram. Nonetheless, much smaller doses of this exotoxin, marketed under various trade names (e.g., Botox), are used to treat muscle spasms and for cosmetic purposes (temporary wrinkle reduction in the skin).
Prokaryotes have amazingly diverse metabolic pathways
Bacteria and archaea outdo the eukaryotes in terms of metabolic diversity. Although they are much more diverse in size and shape, eukaryotes draw on fewer metabolic mechanisms for their energy needs. In fact, much of the eukaryotes’ energy metabolism is carried out in organelles—mitochondria and chloroplasts—that are endosymbiotic descendants of bacteria. The long evolutionary history of bacteria and archaea, during which they have had time to explore a wide variety of habitats, has led to the extraordinary diversity of their metabolic “lifestyles”—their use or nonuse of oxygen, their energy sources, their sources of carbon atoms, and the materials they release as waste products.
Anaerobic Versus Aerobic Metabolism
Some prokaryotes can live only by anaerobic metabolism because oxygen is poisonous to them. These oxygen-sensitive organisms are called obligate anaerobes. Other prokaryotes can shift their metabolism between anaerobic and aerobic modes and are thus called facultative anaerobes. Many facultative anaerobes alternate between anaerobic metabolism (such as fermentation) and cellular respiration as conditions dictate. Aerotolerant anaerobes cannot conduct aerobic cellular respiration, but they are not damaged by oxygen when it is present. By definition, an anaerobe does not use oxygen as an electron acceptor for its respiration.
At the other extreme from the obligate anaerobes, some prokaryotes are obligate aerobes, unable to survive for extended periods in the absence of oxygen. They require oxygen for cellular respiration.
Nutritional Categories
All living organisms face the same nutritional challenges: they must synthesize energy-rich compounds such as ATP to power their life-sustaining metabolic reactions, and they must obtain carbon atoms to build their own organic molecules. Biologists recognize four broad nutritional categories of organisms: photoautotrophs, photoheterotrophs, chemoautotrophs, and chemoheterotrophs. Prokaryotes are represented in all four groups (TABLE 19.2).
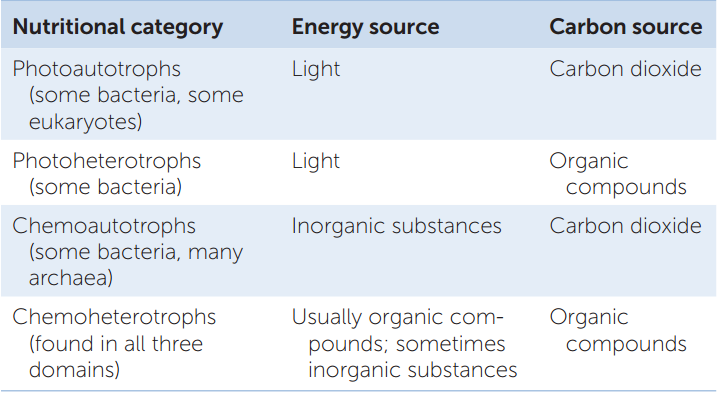
Photoautotrophs perform photosynthesis. They use light as their energy source and carbon dioxide (CO2) as their carbon source. The cyanobacteria, like green plants and other photosynthetic eukaryotes, use chlorophyll a as their key photosynthetic pigment and produce oxygen gas (O2) as a by-product of noncyclic electron transport.
There are other photoautotrophs among the bacteria, but these organisms use bacteriochlorophyll as their key photosynthetic pigment, and they do not produce O2. Instead, some of these photosynthesizers produce particles of pure sulfur, because hydrogen sulfide (H2S), rather than H2O, is their electron donor for photophosphorylation. Many proteobacteria fit into this category. Bacteriochlorophyll molecules absorb light of longer wavelengths than the chlorophyll molecules used by other photosynthesizing organisms. As a result, bacteria using this pigment can grow in water under fairly dense layers of algae, using light of wavelengths that are not absorbed by the algae (FIGURE 19.22).
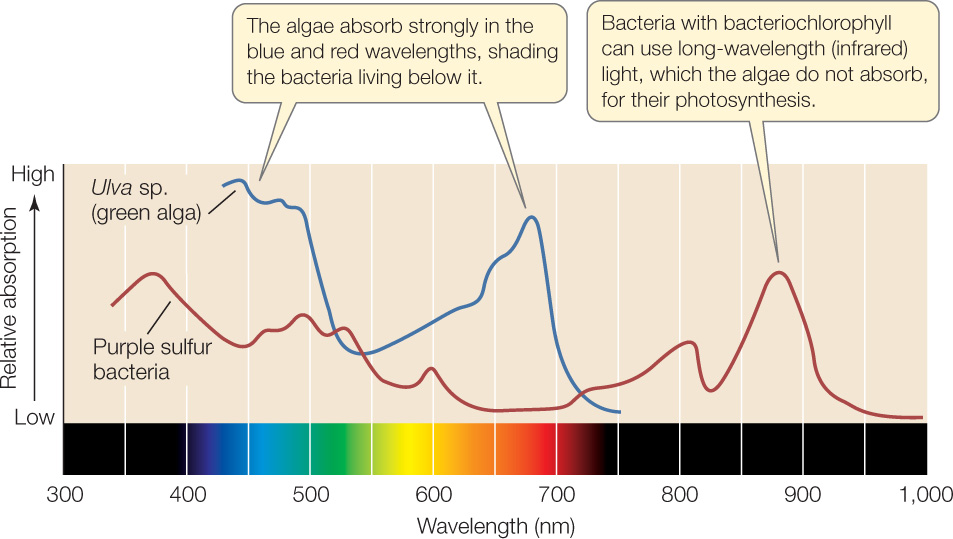
Photoheterotrophs use light as their energy source but must obtain their carbon atoms from organic compounds made by other organisms. Their “food” consists of organic compounds such as carbohydrates, fatty acids, and alcohols. For example, compounds released from plant roots (as in rice paddies) or from decomposing photosynthetic bacteria in hot springs are taken up by photoheterotrophs and metabolized to form building blocks for other compounds. Sunlight provides the energy necessary for metabolism through photophosphorylation.
395
Chemoautotrophs obtain their energy by oxidizing inorganic substances, and they use some of that energy to fix carbon. Some chemoautotrophs use reactions identical to those of the typical photosynthetic cycle, but others use alternative pathways for carbon fixation. Some bacteria oxidize ammonia or nitrite ions to form nitrate ions. Others oxidize hydrogen gas, hydrogen sulfide, sulfur, and other materials. Many archaea are chemoautotrophs.
Finally, chemoheterotrophs obtain both energy and carbon atoms from one or more complex organic compounds that have been synthesized by other organisms. Most known bacteria and archaea are chemoheterotrophs—as are all animals and fungi and many protists.
Although most chemoheterotrophs rely on the breakdown of organic compounds for energy, some chemoheterotrophic prokaryotes obtain their energy by breaking down inorganic substances. Organisms that obtain energy from oxidizing inorganic substances (both chemoautotrophs as well as some chemoheterotrophs) are also known as lithotrophs (Greek, “rock consumers”).
Prokaryotes play important roles in element cycling
The metabolic diversity of the prokaryotes makes them key players in the cycles that keep elements moving through ecosystems. Many prokaryotes are decomposers: organisms that metabolize organic compounds from dead organic material and return the products to the environment as inorganic substances. Prokaryotes, along with fungi, return tremendous quantities of carbon to the atmosphere as carbon dioxide, thus carrying out a key step in the carbon cycle.
The key metabolic reactions of many prokaryotes involve nitrogen or sulfur. For example, some bacteria carry out respiratory electron transport without using oxygen as an electron acceptor. These organisms use oxidized inorganic ions such as nitrate, nitrite, or sulfate as electron acceptors. Examples include the denitrifiers, which release nitrogen to the atmosphere as nitrogen gas (N2). These normally aerobic bacteria, mostly species of the genera Bacillus and Pseudomonas, use nitrate (NO3−) as an electron acceptor in place of oxygen if they are kept under anaerobic conditions:
2 NO3− + 10 e− + 12 H+ → N2 + 6 H2O
Denitrifiers play a key role in the cycling of nitrogen through ecosystems. Without denitrifiers, which convert nitrate ions back into nitrogen gas, all forms of nitrogen would leach from the soil and end up in lakes and oceans, making life on land much more difficult.
Nitrogen fixers convert atmospheric nitrogen gas into a chemical form (ammonia) that is usable by the nitrogen fixers themselves as well as by other organisms:
N2 + 6 H → 2 NH3
All organisms require nitrogen in order to build proteins, nucleic acids, and other important compounds. Nitrogen fixation is thus vital to life as we know it. This all-important biochemical process is carried out by a wide variety of archaea and bacteria (including cyanobacteria) but by no other organisms, so we depend on these prokaryotes for our very existence.
LINK
For descriptions of the role of nitrogen in plant nutrition and of the global nitrogen cycle, see Concepts 25.2 and 45.3
Ammonia is oxidized to nitrate in soil and in seawater by chemoautotrophic bacteria called nitrifiers. Bacteria of two genera, Nitrosomonas and Nitrosococcus, convert ammonia (NH3) to nitrite ions (NO2−), and Nitrobacter oxidize nitrite to nitrate (NO3−), the form of nitrogen most easily used by many plants. What do the nitrifiers get out of these reactions? Their metabolism is powered by the energy released by the oxidation of ammonia or nitrite. For example, by passing the electrons from nitrite through an electron transport system, Nitrobacter can make ATP and, using some of this ATP, can also make NADH. With this ATP and NADH, the bacterium can convert CO2 and H2O into glucose.
396
We have already seen the importance of the cyanobacteria in the cycling of oxygen: in ancient times, the oxygen generated by their photosynthesis converted Earth’s atmosphere from an anaerobic to an aerobic environment. Other prokaryotes—both bacteria and archaea—contribute to the cycling of sulfur. Deep-sea hydrothermal vent ecosystems depend on chemoautotrophic prokaryotes that are incorporated into large communities of crabs, mollusks, and giant worms, all living at a depth of 2,500 meters—below any hint of sunlight. These bacteria obtain energy by oxidizing hydrogen sulfide and other substances released in the near-boiling water flowing from volcanic vents in the ocean floor.
CHECKpoint CONCEPT 19.3
- How do biofilms form, and why are they of special interest to researchers?
- What are three ways that modern lifestyles may be disrupting human microbiomes in ways that affect our health?
- How is nitrogen metabolism in the prokaryotes vital to other organisms? Given the roles of bacteria in the nitrogen cycle, how would you answer people who consider all bacteria to be “germs” and dangerous?
Before moving on to discuss the diversity of eukaryotic life, it is appropriate to consider another category of life that includes some pathogens: the viruses. Although they are not cellular, viruses are numerically among the most abundant forms of life on Earth. Their effects on other organisms are enormous. Where did viruses come from, and how do they fit into the tree of life? Biologists are still working to answer these questions.