CONCEPT21.3 Vascular Tissues Led to Rapid Diversification of Land Plants
The first plants possessing vascular tissues did not arise until tens of millions of years after the earliest plants had colonized the land. But once vascular tissues arose, their ability to transport water and food throughout the plant body allowed vascular plants to spread to new terrestrial environments and to diversify rapidly.
Vascular tissues transport water and dissolved materials
Vascular plants differ from the other land plants in crucial ways, one of which is the possession of a well-developed vascular system consisting of tissues that are specialized for the transport of materials from one part of the plant to another. One type of vascular tissue, the xylem, conducts water and minerals from the soil to aerial parts of the plant. Because some of its cell walls contain a stiffening substance called lignin, xylem also provides support against gravity in the terrestrial environment. The other type of vascular tissue, the phloem, conducts the products of photosynthesis from sites where they are produced or released to sites where they are used or stored.
LINK
The vascular tissues of plants are described in detail in Chapter 24
Although the vascular plants are an extraordinarily large and diverse group, a particular event was critical to their evolution. Sometime during the Paleozoic era, probably in the mid-Silurian (430 mya), a new cell type—the tracheid—evolved in sporophytes of the earliest vascular plants. The tracheid is the principal water-conducting element of the xylem in all vascular plants except the angiosperms (flowering plants) and one small group of gymnosperms—and tracheids persist even in these groups, along with a more specialized and efficient system derived from them.
The evolution of tracheids set the stage for the complete and permanent invasion of land by plants. First, these cells provided a pathway for the transport of water and mineral nutrients from a source of supply to regions of need in the plant body. And second, the cell walls of tracheids, stiffened by lignin, provided rigid structural support. This support is a crucial factor in a terrestrial environment because it allows plants to grow upward and thus compete for sunlight. A taller plant can intercept more direct sunlight (and thus conduct photosynthesis more readily) than a shorter plant, which may be shaded by the taller one. Increased height also improves the dispersal of spores.
The vascular plants featured another evolutionary novelty: a branching, independent sporophyte. A branching sporophyte body can produce more spores than an unbranched body, and it can develop in complex ways. The sporophyte of a vascular plant is nutritionally independent of the gametophyte at maturity. Among the vascular plants, the sporophyte is the large and obvious plant that one normally pays attention to in nature, in contrast to the relatively small, dependent sporophytes typical of most nonvascular land plants.
The diversification of vascular plants made land more suitable for animals
The initial absence of herbivores (plant-eating animals) on land helped make the first vascular plants successful. By the late Silurian period (about 425 mya), the proliferation of land plants made the terrestrial environment more hospitable to animals. Arthropods, vertebrates, and other animals moved onto land only after vascular plants became established there.
429
Trees of various kinds appeared in the Devonian period and dominated the landscape of the Carboniferous period (359–299 mya). Forests of lycophytes (club mosses) up to 40 meters tall, along with horsetails and tree ferns, flourished in the tropical swamps of what would become North America and Europe (FIGURE 21.7). Plant parts from those forests sank into the swamps and were gradually covered by layers of sediment. Over millions of years, as the buried plant material was subjected to intense pressure and elevated temperatures, it was transformed into coal. Today that coal provides more than half of our electricity. The world’s coal deposits, although huge, are not infinite, and humans are burning coal deposits at a far faster rate than they were produced.

In the subsequent Permian period, when the continents came together to form Pangaea, the continental interior became warmer and drier. The 200-million-year reign of the lycophyte–fern forests came to an end as they were replaced by forests of early gymnosperms.
The earliest vascular plants lacked roots
The earliest known vascular plants belonged to a now-extinct group called the rhyniophytes. The rhyniophytes were one of a very few types of vascular plants in the Silurian period. The landscape at that time probably consisted mostly of bare ground, with stands of rhyniophytes in low-lying moist areas. Early versions of the structural features of all the other vascular plant groups appeared in the rhyniophytes of that time. These shared features strengthen the case for the origin of all vascular plants from a common nonvascular land plant ancestor.
Rhyniophytes did not have roots. Like most modern ferns and lycophytes, they were apparently anchored in the soil by horizontal portions of stem, called rhizomes, which bore water-absorbing unicellular filaments called rhizoids. These plants also bore aerial branches, and sporangia—homologous to the sporangia of mosses—were found at the tips of those branches. Their branching pattern was dichotomous, meaning the apex (tip) of the shoot divided to produce two equivalent new branches, each pair diverging at approximately the same angle from the original stem.
The lycophytes are sister to the other vascular plants
The club mosses and their relatives, the spike mosses and quillworts, are collectively called lycophytes. The lycophytes are the sister group to the remaining vascular plants (see Figure 21.1B). There are relatively few (just over 1,200) surviving species of lycophytes.
The lycophytes have true roots that branch dichotomously. The arrangement of vascular tissue in their stems is simpler than that in other vascular plants. They bear simple leaflike structures called microphylls, which are arranged spirally on the stem. Growth in lycophytes comes entirely from apical cell division. Branching in the stems, which is also dichotomous, occurs by division of an apical cluster of dividing cells.
The sporangia of many club mosses are aggregated in cone-like structures called strobili (singular strobilus; FIGURE 21.8A). The strobilus of a club moss is a cluster of spore-bearing leaves inserted on an axis (a linear supporting structure). Other club mosses lack strobili and bear their sporangia on (or adjacent to) the upper surfaces of specialized leaves.

Horsetails and ferns constitute a clade
The horsetails and ferns were once thought to be only distantly related. As a result of genomic analyses, we now know that they form a clade: the monilophytes. In the monilophytes—as in the seed plants, to which they are the sister group (see Figure 21.1B)—there is differentiation between a main stem and side branches. This pattern contrasts with the dichotomous branching characteristic of the lycophytes and rhyniophytes, in which each split gives rise to two branches of similar size.
430
Today there are only about 15 species of horsetails, all in the genus Equisetum. The horsetails have reduced true leaves that form in distinct whorls (circles) around the stem (FIGURE 21.8B). Horsetails are sometimes called “scouring rushes” because rough silica deposits found in their cell walls once made them useful for cleaning. They have true roots that branch irregularly. Horsetails have a large sporophyte and a small gametophyte, each independent of the other.
The first ferns appeared during the Devonian period. Today this group comprises more than 12,000 species. Analyses of gene sequences indicate that a few species traditionally allied with ferns may in fact be more closely related to horsetails than to ferns. Nonetheless, the majority of ferns form a monophyletic group.
Although most ferns are terrestrial, a few species live in shallow fresh water (FIGURE 21.8C). Terrestrial ferns are characterized by large leaves with branching vascular strands (FIGURE 21.8D). Some fern leaves become climbing organs and may grow to be as long as 30 meters.
In the alternating generations of a fern, the gametophyte is small, delicate, and short-lived, but the sporophyte can be very large and can sometimes survive for hundreds of years (FIGURE 21.9). Ferns require liquid water for the transport of the male gametes to the female gametes, so most ferns inhabit shaded, moist woodlands and swamps. The sporangia of ferns typically are borne on a stalk in clusters called sori (singular sorus). The sori are found on the undersurfaces of the leaves, sometimes covering the entire undersurface and sometimes located at the edges.
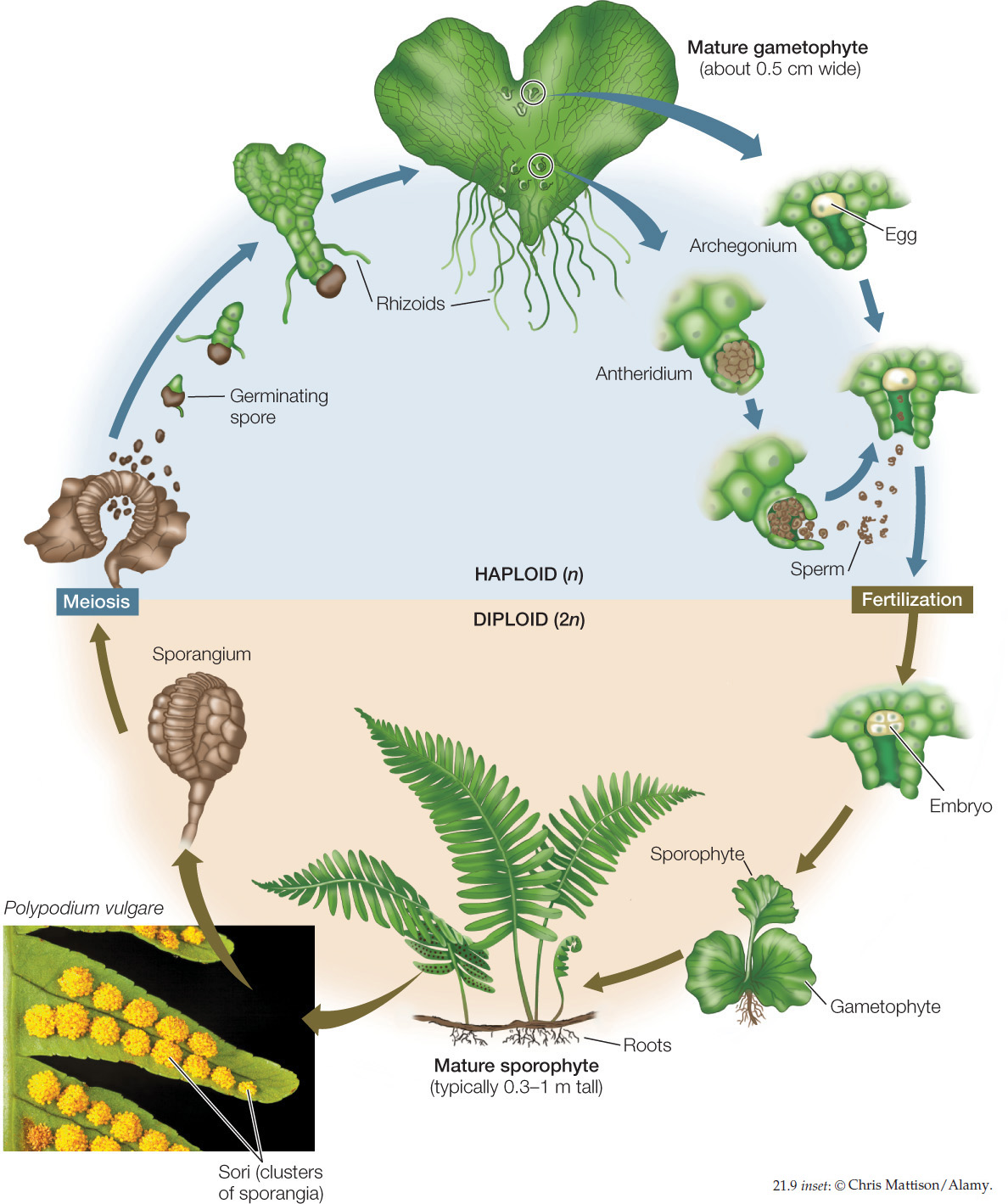
431
The vascular plants branched out
Several features that were new to the vascular plants evolved in lycophytes and monilophytes. Roots probably had their evolutionary origins as a branch, either of a rhizome or of the aboveground portion of a stem. That branch presumably penetrated the soil and branched further. The underground portion could anchor the plant firmly, and even in this primitive condition, it could absorb water and minerals.
The microphylls of lycophytes were probably the first leaflike structures to evolve among the vascular plants. Microphylls are usually small and only rarely have more than a single vascular strand, at least in existing species. Some biologists believe that microphylls had their evolutionary origins as sterile sporangia (FIGURE 21.10A). The principal characteristic of a microphyll is a vascular strand that departs from the vascular system of the stem in such a way that the structure of the stem’s vascular system is scarcely disturbed. This pattern was evident even in the lycophyte trees of the Carboniferous period, many of which had microphylls many centimeters long.
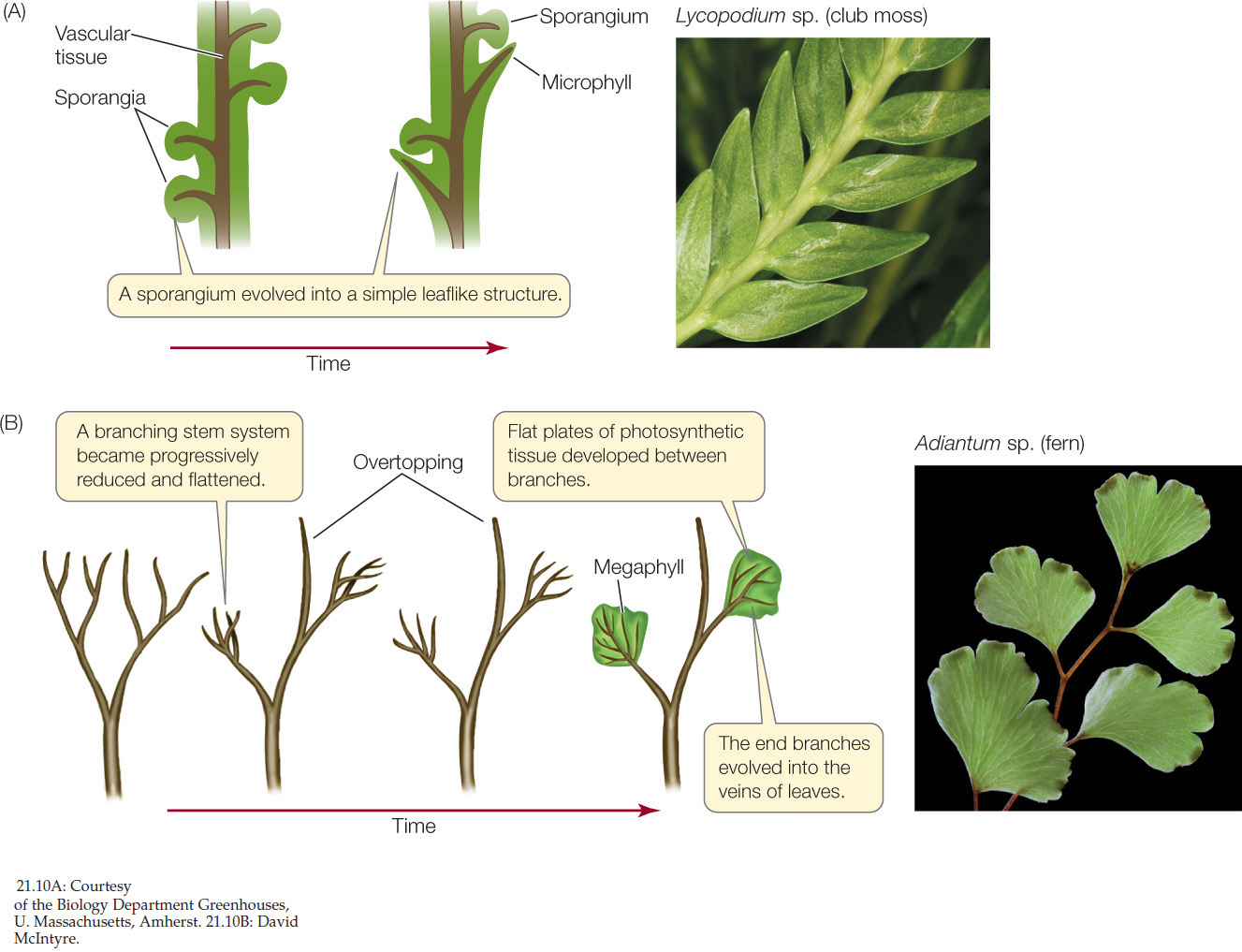
The monilophytes and seed plants constitute a clade called the euphyllophytes (eu, “true”; phyllos, “leaf”). An important synapomorphy of the euphyllophytes is overtopping, a growth pattern in which one branch differentiates from and grows beyond the others (FIGURE 21.10B). Overtopping would have given these plants an advantage in the competition for light, enabling them to shade their dichotomously branching competitors. The overtopping growth of the euphyllophytes also allowed a new type of leaflike structure to evolve. This larger, more complex leaf is called a megaphyll. The megaphyll is thought to have arisen from the flattening of a portion of a branching stem system that exhibited overtopping growth. This change was followed by the development of photosynthetic tissue between the members of overtopped groups of branches, which had the advantage of increasing the photosynthetic surface area of those branches.
Heterospory appeared among the vascular plants
Among the earliest vascular plants, the gametophyte and the sporophyte were independent of one another, and both were photosynthetic. The spores produced by the sporophyte were of a single type and developed into a single type of gametophyte that bore both female and male reproductive organs. This is still true in some surviving lineages of vascular plants, such as horsetails and many ferns. Such plants, which bear a single type of spore, are said to be homosporous (FIGURE 21.11A).
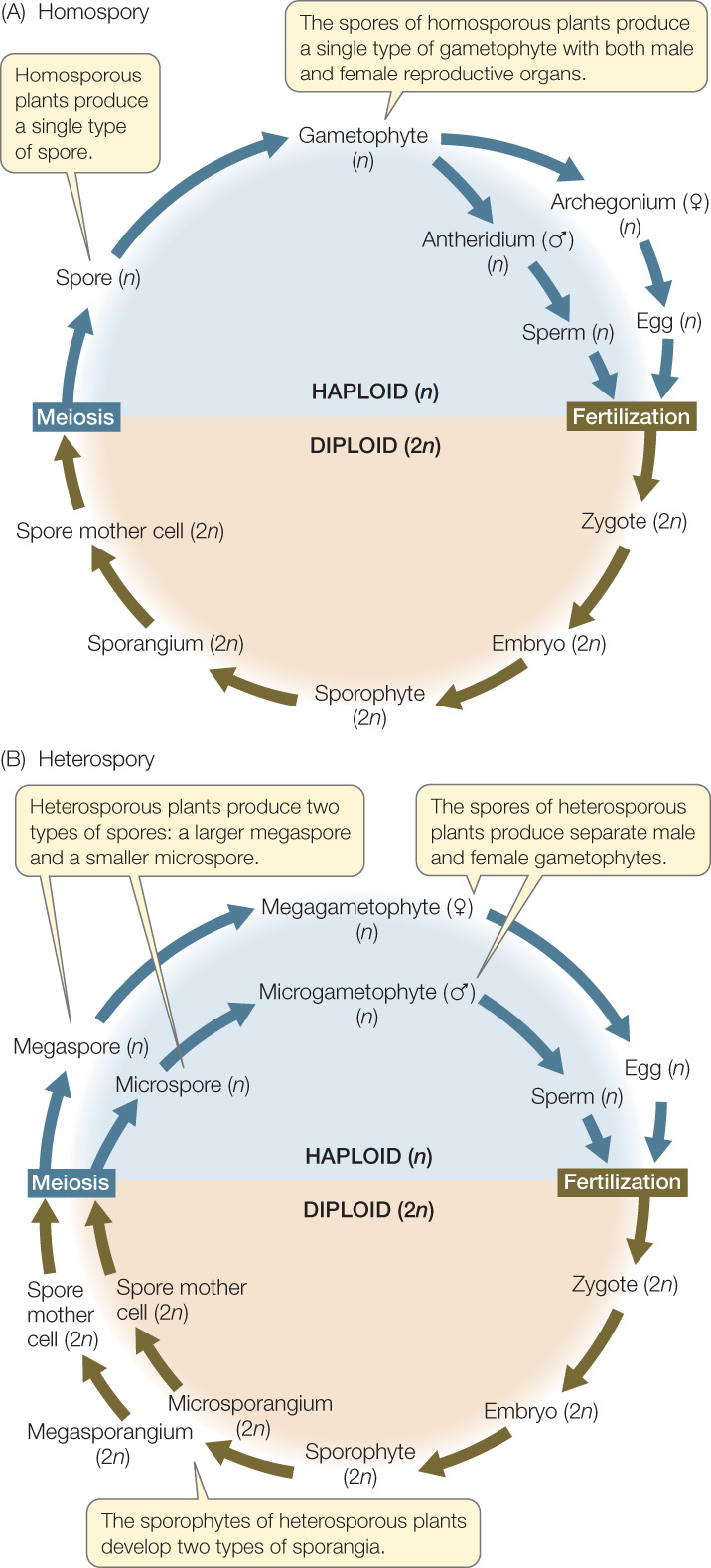
A system with two distinct types of spores evolved several times among the monilophytes and in the ancestor of seed plants. Plants of this type are said to be heterosporous (FIGURE 21.11B). In heterospory, one type of spore—the megaspore—develops into a specifically female gametophyte (a megagametophyte) that produces only eggs. The other type, the microspore, is smaller and develops into a male gametophyte (a microgametophyte) that produces only sperm. The sporophyte produces megaspores in small numbers in megasporangia and microspores in large numbers in microsporangia. Heterospory affects not only the spores and the gametophytes, but also the sporophyte plant itself, which must develop two types of sporangia.
432
The fact that heterospory evolved repeatedly suggests that it affords selective advantages. What advantages does heterospory provide? Heterospory allows the production of many small microspores, which are easily transported from plant to plant. Heterospory also results in the production of a few large megaspores in large megasporangia, which can provide nutrition and protection for the developing embryo. With this division, plants can increase the opportunities for long-distance cross-fertilization, and at the same time increase the chances for survival of the developing embryo.
CHECKpointCONCEPT21.3
- How do the vascular tissues xylem and phloem serve the vascular plants?
- Describe the evolution and distribution of different kinds of leaves and roots among the vascular plants.
- Explain the concept of heterospory. How does heterospory provide selective advantages over homospory?
All of the vascular plant groups we have discussed thus far disperse by means of spores. The embryos of these seedless vascular plants develop directly into sporophytes, which either survive or die, depending on environmental conditions. The spores of some seedless plants may remain dormant and viable for long periods, but the embryos of seedless plants are relatively unprotected (see Figure 21.9). Greater protection of the embryo evolved in the seed plants.