CONCEPT21.5 Flowers and Fruits Increase the Reproductive Success of Angiosperms
The most obvious feature defining the angiosperms is the flower, which is their sexual structure. Production of fruits is also a shared derived trait of angiosperms. After fertilization, the ovary of a flower (together with the seeds it contains) develops into a fruit that protects the seeds and can promote seed dispersal. As we will see, both flowers and fruits afford major reproductive advantages to angiosperms.
Angiosperms have many shared derived traits
The name angiosperm (“enclosed seed”) is derived from another distinctive trait of flowering plants that is related to the formation of fruits: the ovules and seeds are enclosed in a modified leaf called a carpel. Besides protecting the ovules and seeds, the carpel often interacts with incoming pollen to prevent self-fertilization, thus favoring cross-fertilization and increasing genetic diversity.
The female gametophyte of angiosperms is even more reduced than that of gymnosperms, usually consisting of only seven cells. Thus the angiosperms represent the current extreme of the trend we have traced throughout the evolution of vascular plants: the sporophyte generation becomes larger and more independent of the gametophyte, while the gametophyte becomes smaller and more dependent on the sporophyte.
The xylem of most angiosperms is distinguished by the presence of specialized water-transporting cells called vessel elements. These cells are larger in diameter than tracheids and connect with one another without obstruction, allowing easy water movement. A second distinctive cell type in angiosperm xylem is the fiber, which plays an important role in supporting the plant body. Angiosperm phloem possesses another unique cell type, called a companion cell. Like the gymnosperms, woody angiosperms show secondary growth, increasing in diameter by producing secondary xylem and secondary phloem.
A more comprehensive list of angiosperm synapomorphies, then, includes the following (some of these traits will be discussed later in this chapter):
- Flowers
- Fruits
- Ovules and seeds enclosed in a carpel
- Highly reduced gametophytes
- Germination of pollen on a stigma
- Double fertilization
- Endosperm (nutritive tissue for the embryo)
- Phloem with companion cells
The majority of these traits bear directly on angiosperm reproduction, which is a large factor in the success of this dominant plant group.
The sexual structures of angiosperms are flowers
Flowers come in an astonishing variety of forms—just think of some of the flowers you recognize. Flowers may be single, or they may be grouped together to form an inflorescence. Different families of flowering plants have characteristic types of inflorescences, such as the compound umbels of the carrot family (FIGURE 21.19A), the heads of the aster family (FIGURE 21.19B), and the spikes of many grasses (FIGURE 21.19C).

If you examine any familiar flower, you will notice that the outer parts look somewhat like leaves. In fact, all the parts of a flower are modified leaves. The diagram in Figure 21.14B represents a generalized flower (for which there is no exact counterpart in nature). The structures bearing microsporangia are called stamens. Each stamen is composed of a filament bearing an anther that contains the pollen-producing microsporangia. The structures bearing megasporangia are called carpels. The swollen base of the carpel, containing one or more ovules (each containing a megasporangium surrounded by two protective integuments), is called the ovary. The stalk at the top of the carpel is the style, and the terminal surface that receives pollen grains is the stigma. Two or more fused carpels, or a single carpel if only one is present, are also called a pistil.
In addition, many flowers have specialized sterile (non-spore-bearing) leaves. The inner ones are called petals (collectively, the corolla) and the outer ones sepals (collectively, the calyx). The corolla and calyx can be quite showy and often play roles in attracting animal pollinators to the flower. The calyx more commonly protects the immature flower in bud. From base to apex, these floral organs—sepals, petals, stamens, and carpels—are usually positioned in circular arrangements or whorls and attached to a central stalk.
The generalized flower in Figure 21.14B has functional megasporangia and microsporangia. Such flowers are referred to as perfect (or hermaphroditic). Many angiosperms produce two types of flowers, one with only megasporangia and the other with only microsporangia. Consequently, either the stamens or the carpels are nonfunctional or absent in a given flower, and the flower is referred to as imperfect.
Species such as corn or birch, in which both megasporangiate (female) and microsporangiate (male) flowers occur on the same plant, are said to be monoecious (“one-housed”—but, it must be added, one house with separate rooms). Complete separation of imperfect flowers occurs in some other angiosperm species, such as willows and date palms; in these species, an individual plant produces either flowers with stamens or flowers with carpels, but never both. Such species are said to be dioecious (“two-housed”).
440
Flower structure has evolved over time
The flowers of the earliest-diverging clades of angiosperms have a large and variable number of tepals (undifferentiated sepals and petals), carpels, and stamens (FIGURE 21.20A). Evolutionary change within the angiosperms has included some striking modifications of this early condition: reductions in the number of each type of floral organ to a fixed number, differentiation of petals from sepals, and changes in symmetry from radial (as in a lily or magnolia) to bilateral (as in a sweet pea or orchid), often accompanied by an extensive fusion of parts (FIGURE 21.20B).

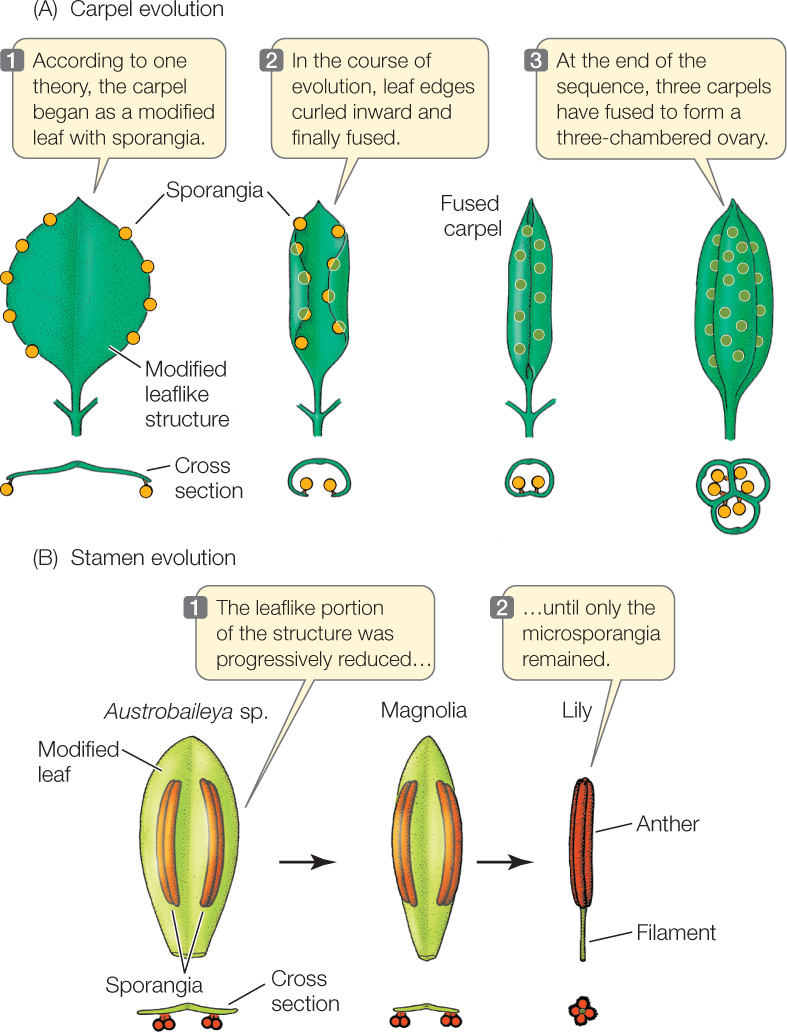
According to one hypothesis, the first carpels to evolve were leaves with marginal sporangia, folded but incompletely closed. Early in angiosperm evolution, the carpels fused with one another, forming a single, multichambered ovary (FIGURE 21.21A). In some flowers, the other floral organs are attached at the top of the ovary, rather than at the bottom as in Figure 21.14B. The stamens of the most ancient flowers may have been leaflike (FIGURE 21.21B), with little resemblance to the stamens of the generalized flower in Figure 21.14B.
A perfect flower represents a compromise of sorts. On the one hand, by attracting a pollinating bird or insect, the plant is attending to both its female and male functions with a single flower type, whereas plants with imperfect flowers must create that attraction twice—once for each type of flower. On the other hand, the perfect flower can favor self-pollination, which is usually disadvantageous. Another potential problem is that the female and male functions might interfere with each other—for example, the stigma might be so placed as to make it difficult for pollinators to reach the anthers, thus reducing the export of pollen to other flowers.
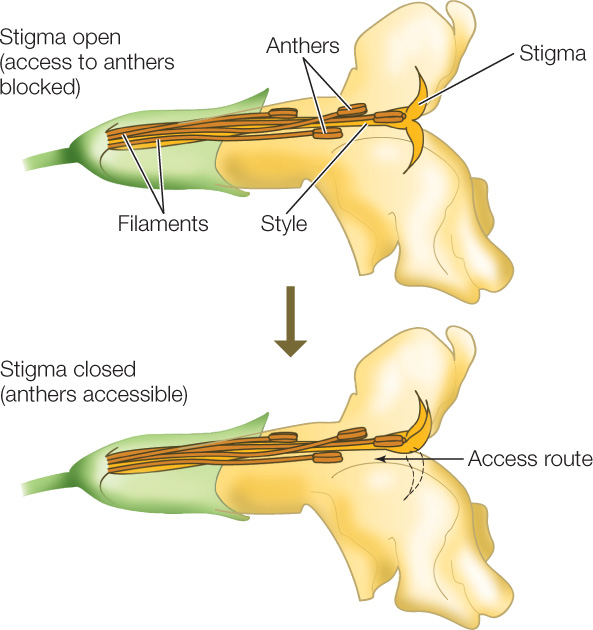
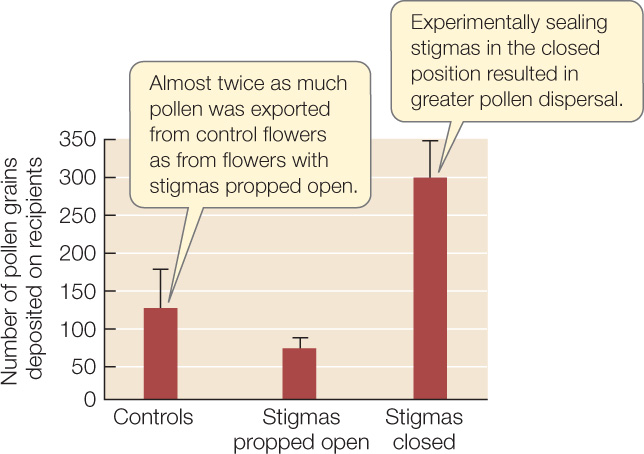
Might there be a way around these problems? One solution is seen in the bush monkeyflower (Mimulus aurantiacus), which is pollinated by hummingbirds. Its flower has a stigma that initially serves as a screen, hiding the anthers (FIGURE 21.22). Once a hummingbird touches the stigma, one of the stigma’s two lobes is retracted, so that subsequent hummingbird visitors pick up pollen from the previously screened anthers. Thus the first bird to visit the flower transfers pollen from another plant to the stigma, eventually leading to fertilization. Later visitors pick up pollen from the now-accessible anthers, fulfilling the flower’s male function. FIGURE 21.23 describes the experiment that revealed the function of this mechanism.
Investigation
HYPOTHESIS
The stigma-retraction response in M. aurantiacus increases the likelihood than an individual flower’s pollen will be exported to another flower once pollen from another flower has been deposited on its stigma.
- Set up three groups of monkeyflower arrays. Each array consists of one pollen-donor flower and multiple pollen-recipient flowers (with the anthers removed to prevent pollen donation).
- In control arrays, the stigma of the pollen donor is allowed to function normally.
- In one set of experimental arrays, the stigma of the pollen donor is permanently propped open.
- In a second set of experimental arrays, the stigma of the pollen donor is artificially sealed closed.
- Allow hummingbirds to visit the arrays, then count the pollen grains transferred from each donor flower to the recipient flowers in the same array.
Error bars show 1 standard error of the mean; the three groups are significantly different at P < 0.05 (see Appendix B).
CONCLUSION
The stigma-retraction response enhances the male function of the flower (dispersal of pollen) once the female function (receipt of pollen) has been performed.
aA. E. Fetscher. 2001. Proceedings of the Royal Society B 268: 525–529.
441
Angiosperms have coevolved with animals
Whereas most gymnosperms are pollinated by wind, most angiosperms are pollinated by animals. The many different mutualistic pollination relationships between plants and animals are vital to both parties. We mentioned coevolution between insects and orchids at the opening of this chapter, but we’ll consider a few additional aspects of plant–pollinator coevolution here.
Many flowers entice animals to visit them by providing food rewards. Pollen grains themselves sometimes serve as food for animals. In addition, some flowers produce a sugary fluid called nectar as a pollinator attractant, and some of these flowers have specialized structures to store and distribute it, as we saw at the opening of this chapter. In the process of visiting flowers to obtain nectar or pollen, animals often carry pollen from one flower to another or from one plant to another. Thus, in their quest for food, the animals contribute to the genetic diversity of the plant population. Insects, especially bees, are among the most important pollinators. Other major pollinators include some species of birds and bats.
For more than 150 million years, angiosperms and their animal pollinators have coevolved in the terrestrial environment. The animals have affected the evolution of the plants, and the plants have affected the evolution of the animals. Flower structure has become incredibly diverse under these selection pressures. Some of the products of coevolution are highly specific. For example, the flowers of some yucca species are pollinated by only one species of yucca moth, and that moth may exclusively pollinate just one species of yucca. Such specific relationships provide plants with a reliable mechanism for transferring pollen only to members of their own species.
Most plant–pollinator interactions are much less specific. In most cases, many different animal species pollinate the same plant species, and the same animal species pollinates many different plant species. However, even these less specific interactions have developed some specialization. Bird-pollinated flowers are often red and odorless. Many insect-pollinated flowers have characteristic odors, and bee-pollinated flowers may have conspicuous markings, called nectar guides, that are conspicuous only to animals, such as bees, that can see colors in the ultraviolet region of the spectrum (FIGURE 21.24).

442
The angiosperm life cycle produces diploid zygotes nourished by triploid endosperms
Like all seed plants, angiosperms are heterosporous. As we have seen, their ovules are contained within carpels rather than being exposed on the surfaces of scales, as in most gymnosperms. The male gametophytes, as in the gymnosperms, are pollen grains.
Pollination in the angiosperms consists of the arrival of a microgametophyte—a pollen grain—on a receptive surface in a flower (the stigma). As in the gymnosperms, pollination is the first in a series of events that results in the formation of a seed. The next event is the growth of a pollen tube extending to the megagametophyte. The third event is a fertilization process that, in detail, is unique to the angiosperms (FIGURE 21.25).
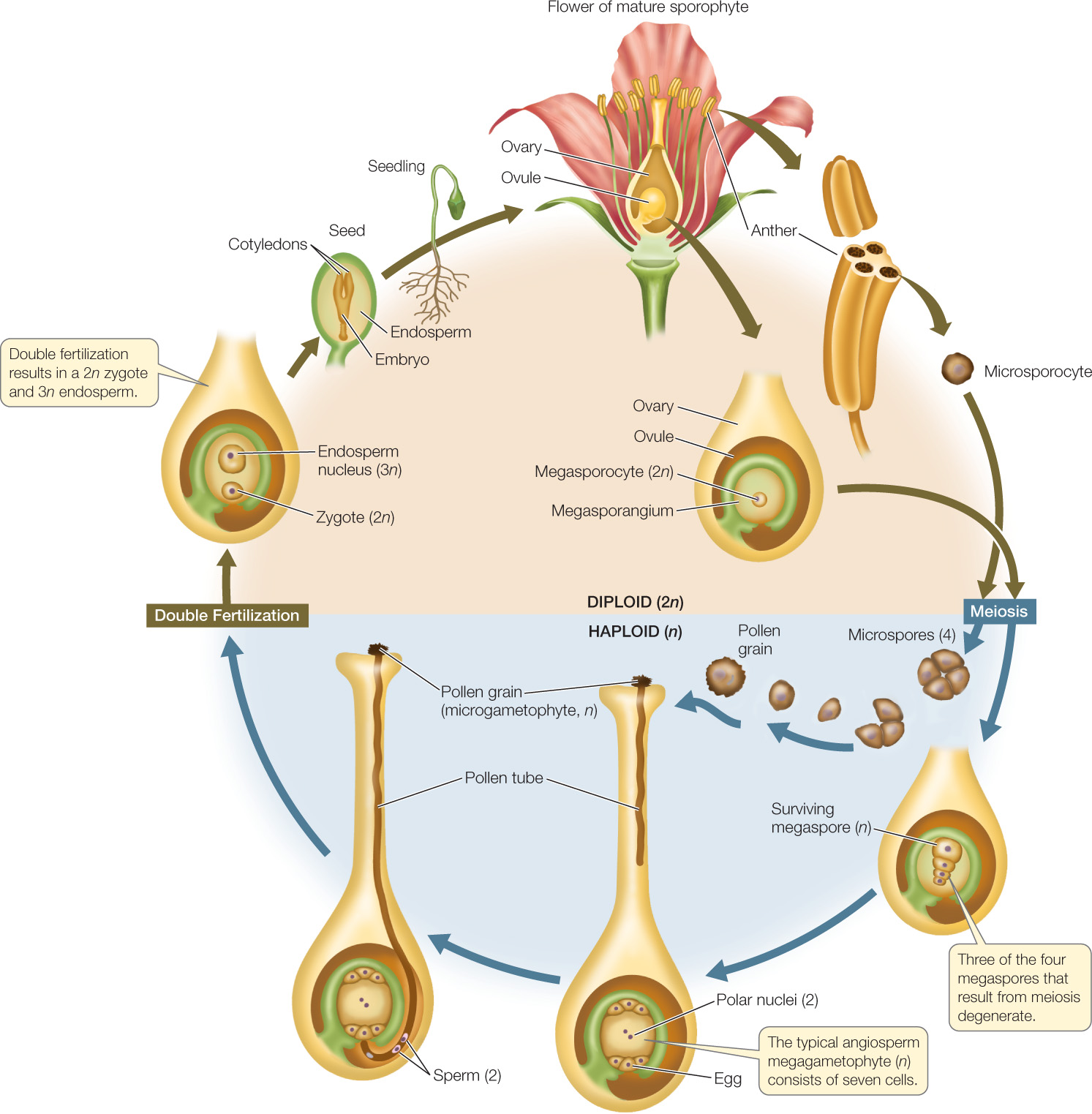
In nearly all angiosperms, two male gametes, contained in a single microgametophyte, participate in fertilization. The nucleus of one sperm combines with that of the egg to produce a diploid zygote, the first cell of the sporophyte generation. In most angiosperms, the other sperm nucleus combines with two other haploid nuclei of the female gametophyte to form a cell with a triploid (3n) nucleus. That cell, in turn, gives rise to triploid tissue, the endosperm, which nourishes the embryonic sporophyte during its early development. This process, in which two fertilization events take place, is known as double fertilization.
443
As Figure 21.25 shows, the zygote develops into an embryo, which consists of an embryonic axis (the “backbone” that will become a stem and a root) and one or two cotyledons, or “seed leaves.” The cotyledons have different fates in different plants. In many, they serve as absorptive organs that take up and digest the endosperm. In others, they enlarge and become photo-synthetic when the seed germinates. Often they play both roles.
The ovule develops into a seed containing the products of the double fertilization that characterizes angiosperms: a diploid zygote and a triploid endosperm (see Figure 21.25). The endosperm serves as storage tissue for starch or lipids, proteins, and other substances that will be needed by the developing embryo.
Fruits aid angiosperm seed dispersal
Fruits typically aid in seed dispersal. Fruits may attach to or be eaten by an animal. The animal is then likely to move, after which the seeds may fall off or be defecated. Fruits are not necessarily fleshy. Fruits can also be hard and woody, or small and have modified structures that allow the seeds to be dispersed by wind or water.
A fruit may consist of only the mature ovary and its seeds, or it may include other parts of the flower or structures associated with it. A simple fruit is one that develops from a single carpel or several fused carpels, such as a plum or peach. A raspberry is an example of an aggregate fruit—one that develops from several separate carpels of a single flower. Pineapples and figs are examples of multiple fruits, formed from a cluster of flowers (an inflorescence). Fruits derived from parts in addition to the carpel and seeds are called accessory fruits—examples are apples, pears, and strawberries (FIGURE 21.26).

444
Fruits increase the reproductive success of angiosperms
Many fleshy fruits attract animals, which eat the fruit and then disperse the seeds in their feces. If all other factors are equal, large seeds have a better chance of producing a successful seedling than small seeds. So why isn’t there selection for larger seeds in the fruits of all plants?
In one study in Peru, the feces of the spider monkey Ateles paniscus were found to contain seeds from 71 species of plants. After eating fruit, the monkeys usually travel some distance before defecating, thus dispersing any undigested seeds.
If monkey feces are left undisturbed on the forest floor, rodents eat and destroy the vast majority of the seeds in the feces. To germinate successfully, the seeds in spider monkey feces need to be buried by dung beetles, which makes the discovery and destruction of seeds by rodents much less likely.
Ellen Andresen hypothesized that dung beetles were more likely to remove larger than smaller seeds from spider monkey dung before burying the dung. She added plastic beads of various diameters to spider monkey dung (to simulate seeds) and measured the percentage of beads buried with the dung by the beetles. Use her data to answer the questions at right.a
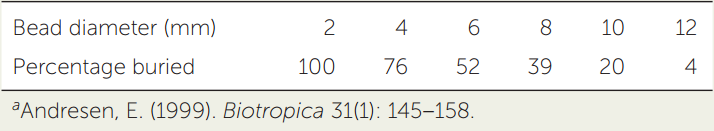
- Plot bead size (the independent variable) versus percentage of beads buried by dung beetles (the dependent variable).
- Calculate a regression line for the relationship shown in your graph (see Appendix B). Approximately what percentage of beads with a diameter of 5 mm would you predict would be buried by the beetles? What about beads 14 mm in diameter?
- What other factors besides size might influence the probability of seed burial by dung beetles? Can you design an experiment to test your hypotheses?
- Describe how changes in the population sizes of spider monkeys, rodents, and dung beetles would affect the reproductive success of various plant species.
445
Recent analyses have revealed the phylogenetic relationships of angiosperms
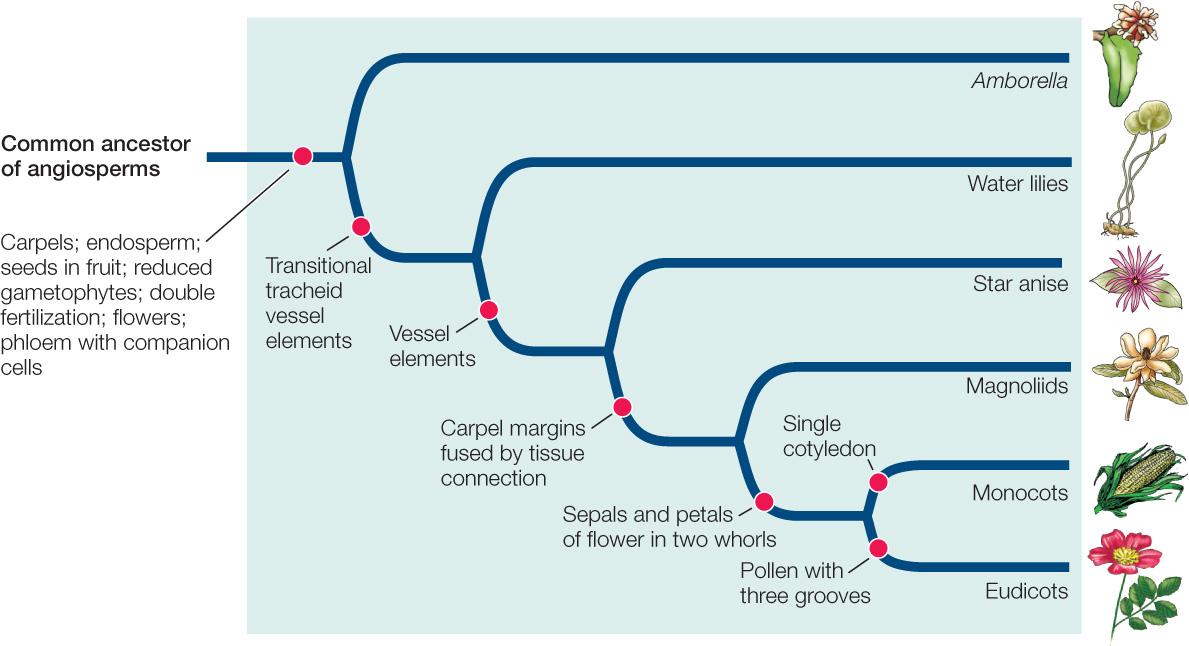
FIGURE 21.27 shows the relationships among the major angiosperm clades. Recent molecular and morphological analyses have supported the hypothesis that the sister group of remaining flowering plants is a single species of the genus Amborella (see Figure 21.28A). This woody shrub, with cream-colored flowers, lives only on New Caledonia, an island in the South Pacific. Other early branching angiosperm groups include the water lilies, star anise and its relatives, and the magnoliids (FIGURE 21.28). The magnoliids include many familiar and useful plants, such as avocados, cinnamon, black pepper, and magnolias.

446
The two largest clades—the monocots and the eudicots—include the great majority of angiosperm species. The monocots are so called because they have a single embryonic cotyledon, whereas the eudicots have two.
Representatives of the two largest angiosperm clades are everywhere. The monocots (FIGURE 21.29) include grasses, cattails, lilies, orchids, and palms. The eudicots (FIGURE 21.30) include the vast majority of familiar seed plants, including most herbs (i.e., nonwoody plants), vines, trees, and shrubs. Among the eudicots are such diverse plants as oaks, willows, beans, snapdragons, roses, and sunflowers.


CHECKpointCONCEPT21.5
- Explain the difference between pollination and fertilization.
- Give some examples of how animals have affected the evolution of the angiosperms.
- What are the respective roles of the two sperm in double fertilization?
- What are the different functions of flowers, fruits, and seeds?
Once life moved onto land, it was plants that shaped the terrestrial environment. Terrestrial ecosystems could not function without the foods and habitats provided by plants. Plants produce oxygen and remove carbon dioxide from the atmosphere. They play important roles in forming soils and renewing their fertility. Plant roots help hold soil in place, providing protection against erosion by wind and water. Plants also moderate local climates in various ways, such as by increasing humidity, providing shade, and blocking wind. All of these ecosystem services permit a great diversity of fungi and animals to exist on land.
447
What was Darwin’s explanation for the three distinct flowers growing on a single orchid plant?
ANSWER After obtaining specimens of the plant in question and dissecting its flowers, Darwin was able to demonstrate that the orchid was a single species (Catasetum macrocarpum) that bore three distinct types of flowers: megasporangiate (female), microsporangiate (male), and perfect (hermaphroditic). The three types of flowers were remarkable in their morphological differences, which had misled botanists into describing the different flower types as species in different genera. Most plants were either male (specimens identified as Catasetum) or female (specimens identified as Monachanthus), but some individuals that bore predominately male or female flowers also produced perfect flowers (specimens identified as Myanthus).
The case of C. macrocarpum demonstrates that some plants blur the lines between strict dioecy (male and female flowers in separate individuals), monoecy (male and female flowers in the same individual), and perfect flowers (flowers with both male and female parts). The flowers on a C. macrocarpum are either male or female at any one time, except that plants can bear some perfect flowers as well.
Why do the male and female flowers of C. macrocarpum look so different? Part of the explanation is their different roles in pollination. Recall Darwin’s observation of a Catasetum flower shooting a packet of pollen at an insect that landed on its flower. The pollinia (pollen packets) and associated structures in male flowers of Catasetum are coiled like springs and are released suddenly when disturbed by an insect. This release forcefully propels the pollinia precisely into position on the back of the insect. The insect pollinator of C. macrocarpum is a specific bee species, the males of which are attracted to the odor of the flowers. The flowers produce no nectar reward, but the male bee does gather the chemical that produces the scent. The bee then moves on to another flower. When the bee visits a female flower on a different C. macrocarpum individual (again attracted by the same scent), no such “loaded spring” awaits. Instead, the morphology of the female flower enhances the removal of the pollinia from the insect’s body. In this way, floral morphology makes cross-fertilization more likely and reduces the chances of self-pollination.
448
Orchids were important in forming Darwin’s ideas about the mechanisms of evolution, for they showed that even aspects of the coloration and form of flowers evolved in response to natural selection. This conclusion ran counter to the thinking of the day. For example, Thomas Huxley, one of Darwin’s earliest and strongest supporters, doubted that the beauty of color in plants and animals could be explained on the basis of their importance to function. Darwin showed that the beauty of flowers is indeed connected to their reproductive success and is a key element in explaining the great diversity of plants.