CONCEPT22.3 Major Groups of Fungi Differ in Their Life Cycles
Major fungal groups were originally defined by their structures and processes for sexual reproduction and also, to a lesser extent, by other morphological differences. Although fungal life cycles are even more diverse than was once realized, specific types of life cycles generally distinguish the six major groups of fungi: microsporidia, chytrids, zygospore fungi (Zygomycota), arbuscular mycorrhizal fungi (Glomeromycota), sac fungi (Ascomycota), and club fungi (Basidiomycota). FIGURE 22.11 diagrams the evolutionary relationships of these groups as they are understood today.
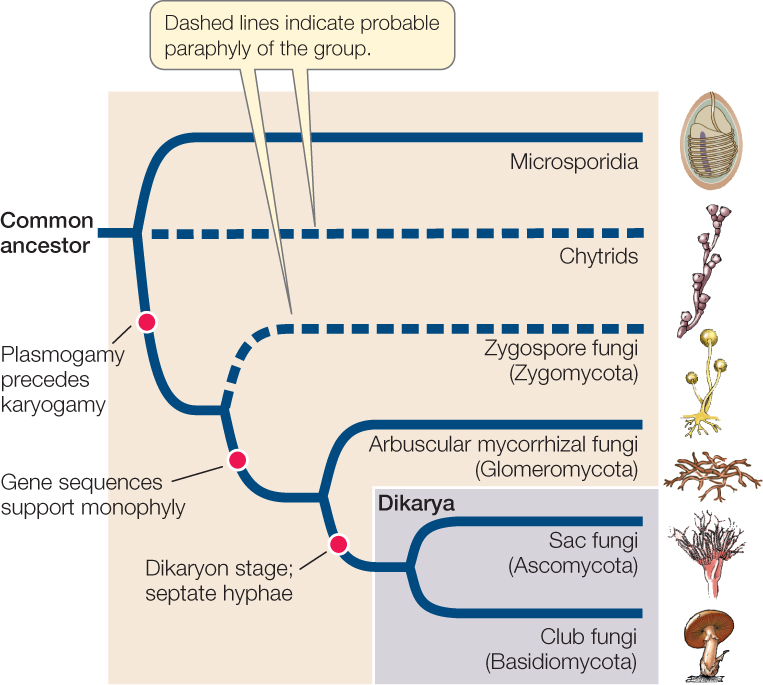
The chytrids and the zygospore fungi may not represent monophyletic groups, as they each consist of several distantly related lineages that retain some ancestral features. The clades that are thought to be monophyletic within these two informal groupings are listed in TABLE 22.1. Recent evidence from DNA analyses has established the placement of the microsporidia among the fungi, the likely paraphyly of the chytrids and the zygospore fungi (in other words, members of these groups are probably not each other’s closest relatives), the independence of arbuscular mycorrhizal fungi from the other fungal groups, and the monophyly of sac fungi and club fungi.
Fungi reproduce both sexually and asexually
Both asexual and sexual reproduction occur among the fungi (FIGURE 22.12). Asexual reproduction takes several forms:
- The production of (usually) haploid spores within sporangia
- The production of haploid spores (not enclosed in sporangia) at the tips of hyphae; such spores are called conidia (Greek konis, “dust”)
- Cell division by unicellular fungi—either a relatively equal division of one cell into two (fission) or an asymmetrical division in which a smaller daughter cell is produced (budding)
- Simple breakage of the mycelium
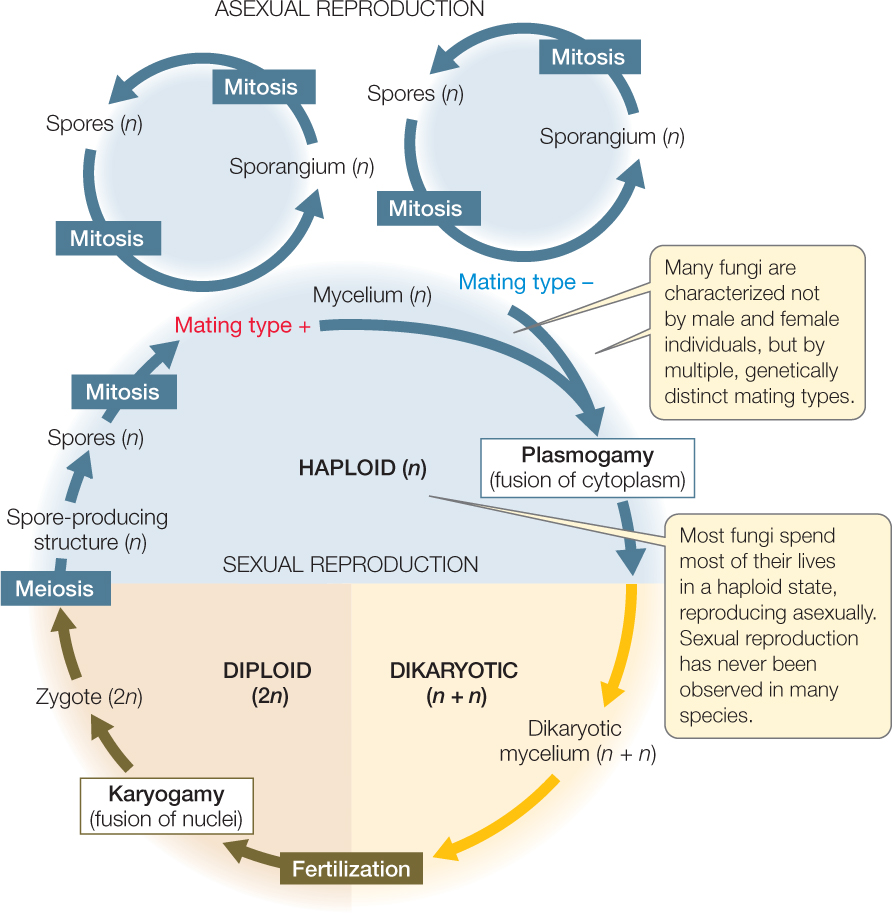
Sexual reproduction is rare (or even unknown) in some groups of fungi but common in others. Sexual reproduction may not occur in some species, or it may occur so rarely that biologists have never observed it. Species in which no sexual stage has been observed were once placed in a separate taxonomic group because knowledge of the sexual life cycle was considered necessary for classifying fungi. Now, however, these species can be related to other species of fungi through analysis of their DNA sequences.
459
Sexual reproduction in most fungi features an interesting twist: There is no morphological distinction between female and male structures, or between female and male individuals, in most groups of fungi. Rather, there is a genetically determined distinction between two or more mating types. Individuals of the same mating type cannot mate with each other, but they can mate with individuals of another mating type within the same species, thus avoiding self-fertilization. Individuals of different mating types differ genetically but are often visually and physiologically indistinguishable.
Microsporidia are highly reduced, parasitic fungi
Microsporidia are unicellular parasitic fungi. They are among the smallest eukaryotes known, with infective spores that are only 1–40 μm in diameter. About 1,500 species have been described, but many more species are thought to exist. Their relationships among the eukaryotes have puzzled biologists for many decades.
Microsporidia lack true mitochondria, although they have reduced structures, known as mitosomes, that are derived from mitochondria. Unlike mitochondria, however, mitosomes contain no DNA—the mitochondrial genome has been completely transferred to the nucleus. Because microsporidia lack mitochondria, biologists initially suspected that they represented an early lineage of eukaryotes that diverged before the endosymbiotic event from which mitochondria evolved. The presence of mitosomes, however, indicates that this hypothesis is incorrect. DNA sequence analysis, along with the fact that their cell walls contain chitin, has confirmed that the microsporidia are in fact highly reduced, parasitic fungi, although their exact placement among the fungal lineages is still being investigated.
460
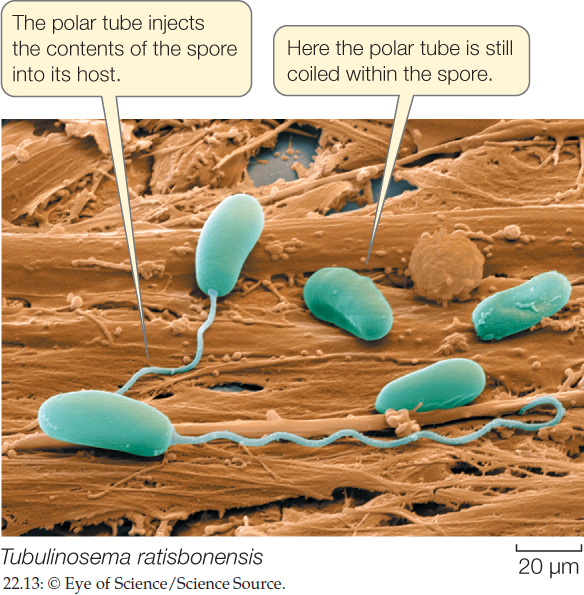
Microsporidia are obligate intracellular parasites of animals, especially of insects, crustaceans, and fishes. Some species are known to infect mammals, including humans. Most infections by microsporidia cause chronic disease in the host, with effects that include weight loss, reduced fertility, and a shortened life span. The host cell is penetrated by a polar tube that grows from the microsporidian spore. The function of the polar tube is to inject the contents of the spore, the sporoplasm, into the host (FIGURE 22.13). The sporoplasm then replicates within the host cell and produces new infective spores. The life cycle of some species is complex and involves multiple hosts, whereas other species infect a single host. In some insects, parasitic microsporidia are transmitted vertically (i.e., from parent to offspring). Reproduction is thought to be strictly asexual in some microsporidians, but includes poorly understood asexual and sexual cycles in other species.
Most chytrids are aquatic
The chytrids include several distinct lineages of mostly aquatic microorganisms once classified with the protists. However, morphological evidence (cell walls that consist primarily of chitin) and molecular evidence support their classification as early-diverging fungi. In this book we use the term “chytrid” to refer to all three of the formally named clades listed as chytrids in Table 22.1, but some mycologists use this term to refer to only one of those clades, the Chytridiomycota. There are fewer than 1,000 described species among the three groups of chytrids.
Chytrids reproduce both sexually and asexually. Like the animals, most chytrids that reproduce sexually possess flagellated gametes. The spores (called zoospores) are also flagellated (FIGURE 22.14A). The retention of flagella in the spores and gametes reflects the aquatic environment in which fungi first evolved. Chytrids are the only fungi that include species with flagella at any life cycle stage.
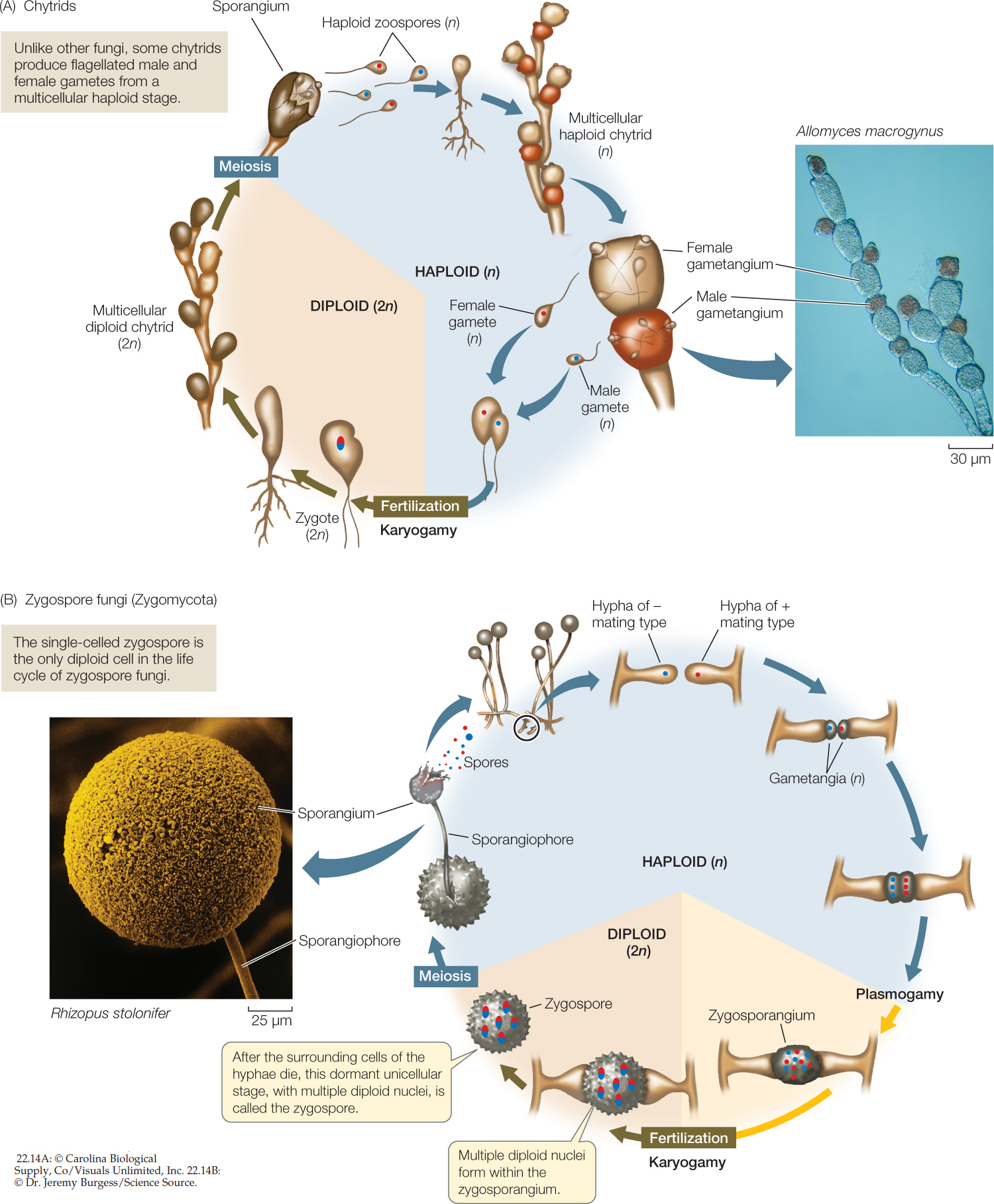
The chytrids are diverse in form. Some are unicellular, others have rhizoids, and still others have coenocytic hyphae. They may be parasitic (on organisms such as algae, mosquito larvae, nematodes, and amphibians) or saprobic. Some have complex mutualistic relationships with foregut-fermenting animals such as cattle and deer. Many chytrids live in freshwater habitats or in moist soil, but some are marine.
Some fungal life cycles feature separate fusion of cytoplasms and nuclei
Most members of the remaining four groups of fungi are terrestrial. Although the terrestrial fungi grow in moist places, they do not have motile gametes, and they do not release gametes into the environment, so liquid water is not required for fertilization. Instead, the cytoplasms of two individuals of different mating types fuse (a process called plasmogamy) before their nuclei fuse (a process called karyogamy). Sexual species of terrestrial fungi include some zygospore fungi, sac fungi, and club fungi.
Zygospore fungi reproduce sexually when adjacent hyphae of two different mating types release chemical signals that cause them to grow toward each other. These hyphae produce gametangia, which are specialized cells for reproduction that are retained as part of the hyphae. In the gametangia, nuclei replicate without cell division, resulting in multiple haploid nuclei in both gametangia. The two gametangia then fuse to form a zygosporangium that contains many haploid nuclei of each mating type (FIGURE 22.14B). Haploid nuclei of different mating types then pair up to form multiple diploid nuclei within the zygosporangium. A thick, multilayered cell wall develops around the zygosporangium to form a well-protected resting stage that can remain dormant for months. In harsh environmental conditions, this resting stage may be the only cell that survives as the surrounding cells of the hyphae die. At this stage the single surviving cell is known as a zygospore, which is the basis of the name of the zygospore fungi. When environmental conditions improve, the nuclei within the zygospore undergo meiosis and one or more stalked sporangiophores sprout, each bearing a sporangium. Each sporangium contains the products of meiosis: haploid nuclei that are incorporated into spores. These spores disperse and germinate to form a new generation of haploid hyphae.
The zygospore fungi include four major lineages of terrestrial fungi that live on soil as saprobes, as parasites of insects and spiders, or as mutualists of other fungi and invertebrate animals. They produce no cells with flagella, and only one diploid cell—the zygospore—appears in the entire life cycle. Their hyphae are coenocytic. Most species do not form a fleshy fruiting structure. Instead, the hyphae spread in a radial pattern from the spore, with occasional stalked sporangiophores reaching up into the air (FIGURE 22.15).
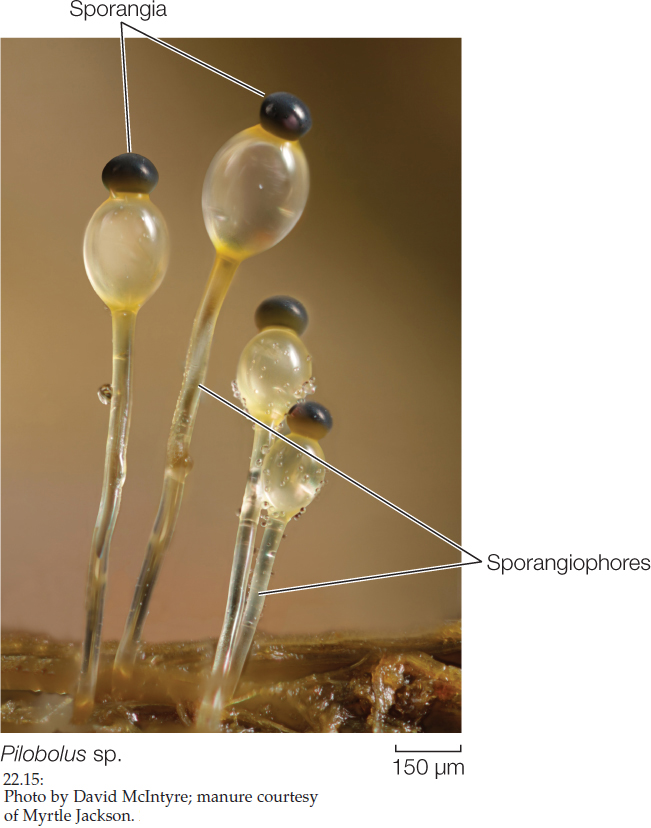
461
462
More than 1,000 species of zygospore fungi have been described. One species you may have seen is Rhizopus stolonifer, the black bread mold. Rhizopus produces many stalked sporangiophores, each bearing a single sporangium containing hundreds of minute spores (see Figure 22.14B).
Arbuscular mycorrhizal fungi form symbioses with plants
Arbuscular mycorrhizal fungi (Glomeromycota) are terrestrial fungi that associate with plant roots in a symbiotic, mutualistic relationship (see Figure 22.10B). Fewer than 200 species have been described, but 80–90 percent of all plants have associations with them. Molecular systematic studies have suggested that arbuscular mycorrhizal fungi are the sister group to the Dikarya (sac fungi and club fungi; see Figure 22.11).
The hyphae of arbuscular mycorrhizal fungi are coenocytic. These fungi use glucose from their plant partners as their primary energy source, converting it into other, fungus-specific sugars that cannot return to the plant. Arbuscular mycorrhizal fungi are only known to reproduce asexually.
The dikaryotic condition is a synapomorphy of sac fungi and club fungi
In the two remaining groups of fungi—the sac fungi and the club fungi—some stages have a nuclear configuration other than the familiar haploid or diploid states (FIGURE 22.16). In these fungi, karyogamy (fusion of nuclei) occurs long after plasmogamy (fusion of cytoplasm), so that two genetically different haploid nuclei coexist and divide within each cell of the mycelium. This stage of the fungal life cycle is called a dikaryon (“two nuclei”) and its ploidy is indicated as n + n. The dikaryon is a synapomorphy of these two groups, which are placed together in a clade called Dikarya.
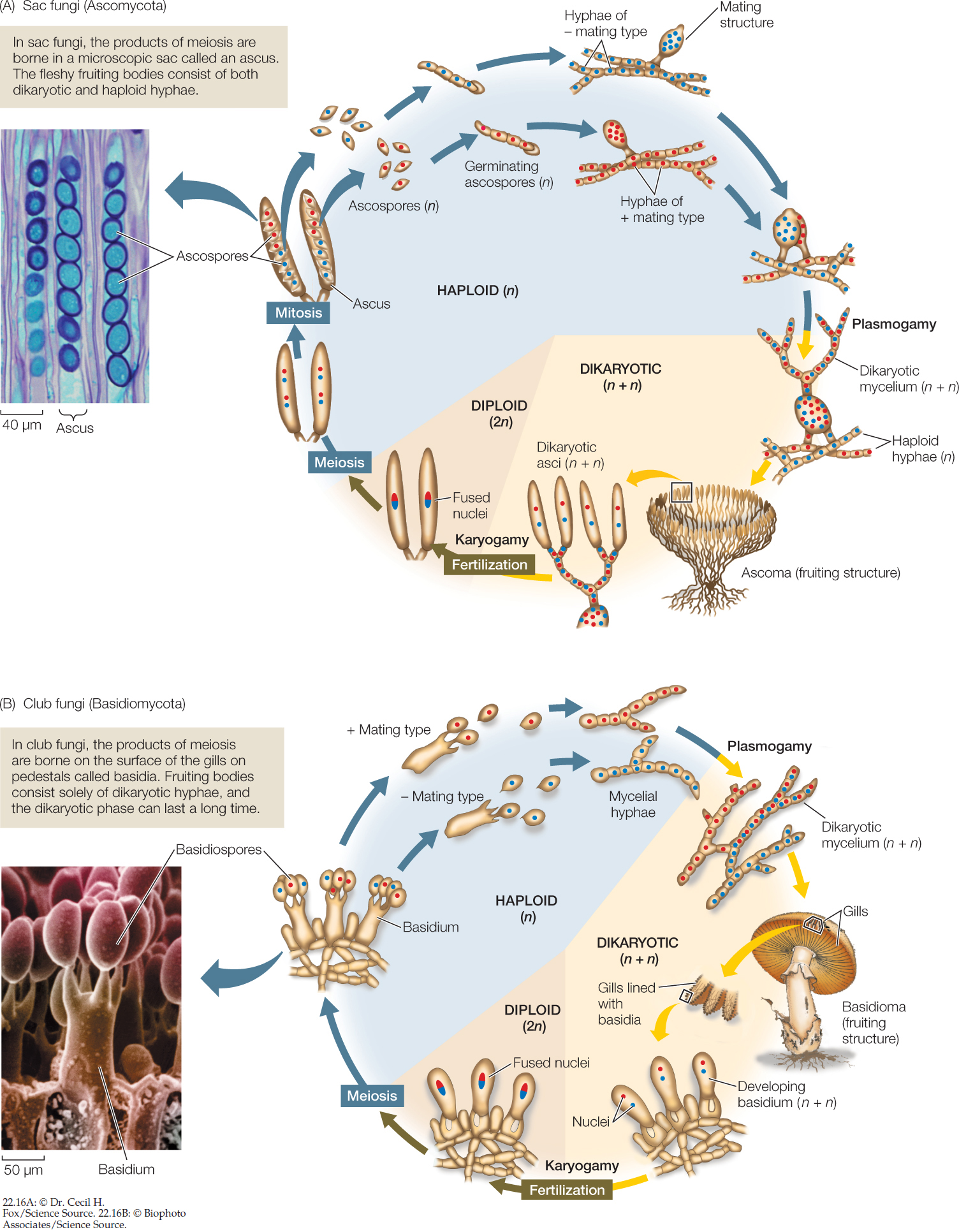
Eventually, specialized fruiting structures form, within which pairs of genetically dissimilar nuclei—one from each parent—fuse, giving rise to zygotes long after the original “mating.” The diploid zygote nucleus undergoes meiosis, producing four haploid nuclei. The mitotic descendants of those nuclei become spores, which germinate to give rise to the next haploid generation.
A life cycle with a dikaryon stage has several unusual features. First, there are no gamete cells, only gamete nuclei. Second, the only true diploid structure is the zygote, although for a long period the genes of both parents are present in the dikaryon and can be expressed. In effect, the dikaryon is neither diploid (2n) nor haploid (n). Rather, it is dikaryotic (n + n). Therefore a harmful recessive mutation in one nucleus may be compensated for by a normal allele on the same chromosome in the other nucleus, and dikaryotic hyphae often have characteristics that are different from their n or 2n products. The dikaryotic condition is perhaps the most distinctive of the genetic peculiarities of the fungi.
The sexual reproductive structure of sac fungi is the ascus
The sac fungi (Ascomycota) are a large and diverse group of fungi found in marine, freshwater, and terrestrial habitats. There are approximately 64,000 known species, almost a third of which are the fungal partners in lichens. The hyphae of sac fungi are segmented by more or less regularly spaced septa. A pore in each septum permits extensive movement of cytoplasm and organelles (including nuclei) from one segment to the next.
Sac fungi are distinguished by the production of sacs called asci (singular ascus), which after meiosis and spore cleavage contain sexually produced haploid ascospores (see Figure 22.16A). The ascus is the characteristic sexual reproductive structure of the sac fungi. In the past, the sac fungi were classified on the basis of whether or not the asci are contained within a specialized fruiting structure known as an ascoma (plural ascomata) and on differences in the morphology of that fruiting structure. DNA sequence analyses have resulted in a revision of these traditional groupings, however.
Sac Fungus Yeasts
Some species of sac fungi are unicellular yeasts. The 1,000 or so species in this group are among the most important domesticated fungi. Perhaps the best known is baker’s, or brewer’s, yeast (Saccharomyces cerevisiae; see Figure 22.2), which metabolizes glucose obtained from its environment into ethanol and carbon dioxide by fermentation. It forms carbon dioxide bubbles in bread dough and gives baked bread its light texture. Although they are baked away in bread making (which produces the pleasant aroma of baking bread), the ethanol and carbon dioxide are both retained when the yeast ferments grain into beer. Other sac fungus yeasts live on fruits such as figs and grapes and play an important role in the making of wine. Many others are associated with insects. In the guts of some insects, they provide enzymes that break down materials that are otherwise difficult for the insects to digest, especially cellulose.
463
464
Sac fungus yeasts reproduce asexually by budding. Sexual reproduction takes place when two adjacent haploid cells of opposite mating types fuse. In some species, the resulting zygote buds to form a diploid cell population. In others, the zygote nucleus undergoes meiosis immediately to form an ascus. Depending on whether the products of meiosis then undergo mitosis, a yeast ascus usually contains either eight or four ascospores, which germinate to become haploid cells. The sac fungus yeasts have lost the dikaryon stage.
Filamentous Sac Fungi
Most sac fungi are filamentous species, such as the cup fungi (FIGURE 22.17A), in which the ascomata are cup-shaped and can be as large as several centimeters across (although most are much smaller). The inner surfaces of the ascomata, which are covered with a mixture of specialized hyphae and asci, produce huge numbers of spores. The edible ascomata of some species, including morels and truffles (FIGURE 22.17B), are regarded by humans as gourmet delicacies (and can sell at prices higher than gold). The underground ascomata of truffles have a strong odor that attracts mammals such as pigs, which then eat and disperse the fungus. Humans sometimes take advantage of pigs’ attraction to truffles and use pigs to locate truffles for human consumption.

The sexual reproductive cycle of filamentous sac fungi includes the formation of a dikaryon, although this stage is relatively brief compared with that in club fungi. Many filamentous sac fungi form multinucleate mating structures (see Figure 22.16A). Mating structures of two different mating types fuse and produce a dikaryotic mycelium, containing nuclei from both mating types. The dikaryotic mycelium typically forms a cup-shaped ascoma, which bears the asci. Only after the formation of asci do the nuclei from the two mating types finally fuse. Both nuclear fusion and the subsequent meiosis that produces haploid ascospores take place within individual asci. The ascospores are ultimately released (sometimes shot off forcefully) by the ascus to begin the new haploid generation.
The sac fungi also include many of the filamentous fungi known as molds. Molds consist of filamentous hyphae that do not form large ascomata, although they can still produce asci and ascospores. Many molds are parasites of flowering plants. Chestnut blight and Dutch elm disease are both caused by molds. The chestnut blight fungus, which was introduced to the United States in the 1890s, had destroyed the American chestnut as a commercial species by 1940. Before the blight, this species accounted for more than half the trees in the eastern North American forests. Another familiar story is that of the American elm. Sometime before 1930, the Dutch elm disease fungus (first discovered in the Netherlands, but native to Asia) was introduced into the United States on infected elm logs from Europe. Spreading rapidly—sometimes by way of connected root systems—the fungus destroyed great numbers of American elm trees.
Other plant pathogens among the sac fungi include the powdery mildews that infect cereal crops, lilacs, and roses, among many other plants. Mildews can be a serious problem to farmers and gardeners, and a great deal of research has focused on ways to control these agricultural pests.
Brown molds of the genus Aspergillus are important in some human diets. A. tamarii acts on soybeans in the production of soy sauce, and A. oryzae is used in brewing the Japanese alcoholic beverage sake. Some species of Aspergillus that grow on grains and on nuts such as peanuts and pecans produce extremely carcinogenic (cancer-inducing) compounds called aflatoxins.
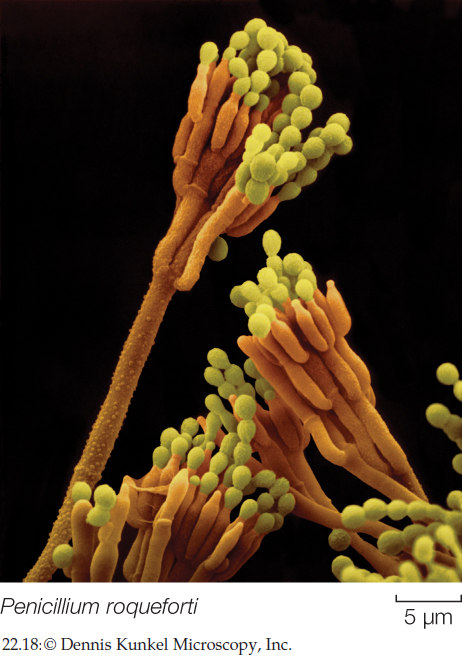
Penicillium is a genus of green molds, of which some species produce the antibiotic penicillin, presumably for defense against competing bacteria. Two species, P. camembertii and P. roqueforti, are the organisms responsible for the characteristic strong flavors of Camembert and Roquefort cheeses, respectively.
The filamentous sac fungi can also reproduce asexually by means of conidia that form at the tips of specialized hyphae (FIGURE 22.18). Small chains of conidia are produced by the millions and can survive for weeks in nature. The conidia are what give molds their characteristic colors.
The sexual reproductive structure of club fungi is the basidium
Club fungi (Basidiomycota) produce some of the most spectacular fruiting structures found among the fungi. These fruiting structures, called basidiomata (singular basidioma), include mushrooms of all kinds, puffballs (see Figure 22.4), and the bracket fungi often encountered on trees and fallen logs in a damp forest. About 30,000 species of club fungi have been described. They include about 4,000 species of mushrooms, including both poisonous and edible species (FIGURE 22.19A). Bracket fungi (FIGURE 22.19B) play an important role in the carbon cycle by breaking down wood. They also do great damage to both cut lumber and timber stands. Some of the most economically damaging plant pathogens are club fungi, including the rust fungi and smut fungi that parasitize cereal grains. In contrast, other club fungi contribute to the survival of plants as fungal partners in ectomycorrhizae.

465
The hyphae of club fungi characteristically have septa with small, distinctive pores. As these hyphae grow, haploid nuclei of different mating types meet and fuse, forming dikaryotic hyphae. The dikaryotic mycelium grows and eventually, when triggered by rain or another environmental cue, produces a basidioma. The dikaryon stage may persist for years, or even centuries. This pattern contrasts with the life cycle of the sac fungi, in which the dikaryon is found only in the stages leading up to formation of the asci.
The basidium (plural basidia), a swollen cell at the tip of a specialized hypha, is the characteristic sexual reproductive structure of the club fungi (see Figure 22.16B). In mushroom-forming club fungi, the basidia typically form on specialized structures known as gills. The basidium is the site of nuclear fusion and meiosis and thus plays the same role in the club fungi as the ascus does in the sac fungi and the zygosporangium does in the zygospore fungi.
After nuclei fuse in the basidium, the resulting diploid nucleus undergoes meiosis, and the four resulting haploid nuclei are incorporated into haploid basidiospores, which form on tiny stalks on the outside of the basidium. A single basidioma of the common bracket fungus Ganoderma applanatum can produce as many as 4.5 trillion basidiospores in one growing season. Basidiospores typically are forcibly discharged from their basidia and then germinate, giving rise to hyphae with haploid nuclei.
CHECKpointCONCEPT22.3
- Explain the concept of mating types. How are they different from male and female sexes?
- Explain how the microsporidia infect the cells of their animal hosts.
- What feature of the chytrids suggests that the fungi had an aquatic ancestor?
- What is the role of the zygospore in the life cycle of zygospore fungi?
- What distinguishes the fruiting structures of sac fungi from those of club fungi?
466
Fungi are of interest to biologists because of the roles they play in interactions with other organisms. But fungi can also be useful as tools for studying environmental problems and for finding solutions to those problems.