CONCEPT23.1 Distinct Body Plans Evolved among the Animals
How do we recognize an animal? The answer may seem obvious for familiar animals, but consider the sponges, which were once thought to be plants, or the rotifers, which are smaller than some protists.
The general characteristics often used to recognize animals include multicellularity, heterotrophy (using nutrients produced by other organisms), internal digestion, and motility (independent movement). Although these features are useful, none is diagnostic for all animals. Some animals do not move independently (at least during certain life stages), and some plants and fungi do have limited movement. Many parasitic animals lack a gut for internal digestion, and many other organisms (notably the fungi) are multicellular and heterotrophic. So what evidence leads us to group the animals in a single clade?
Animal monophyly is supported by gene sequences and cellular morphology
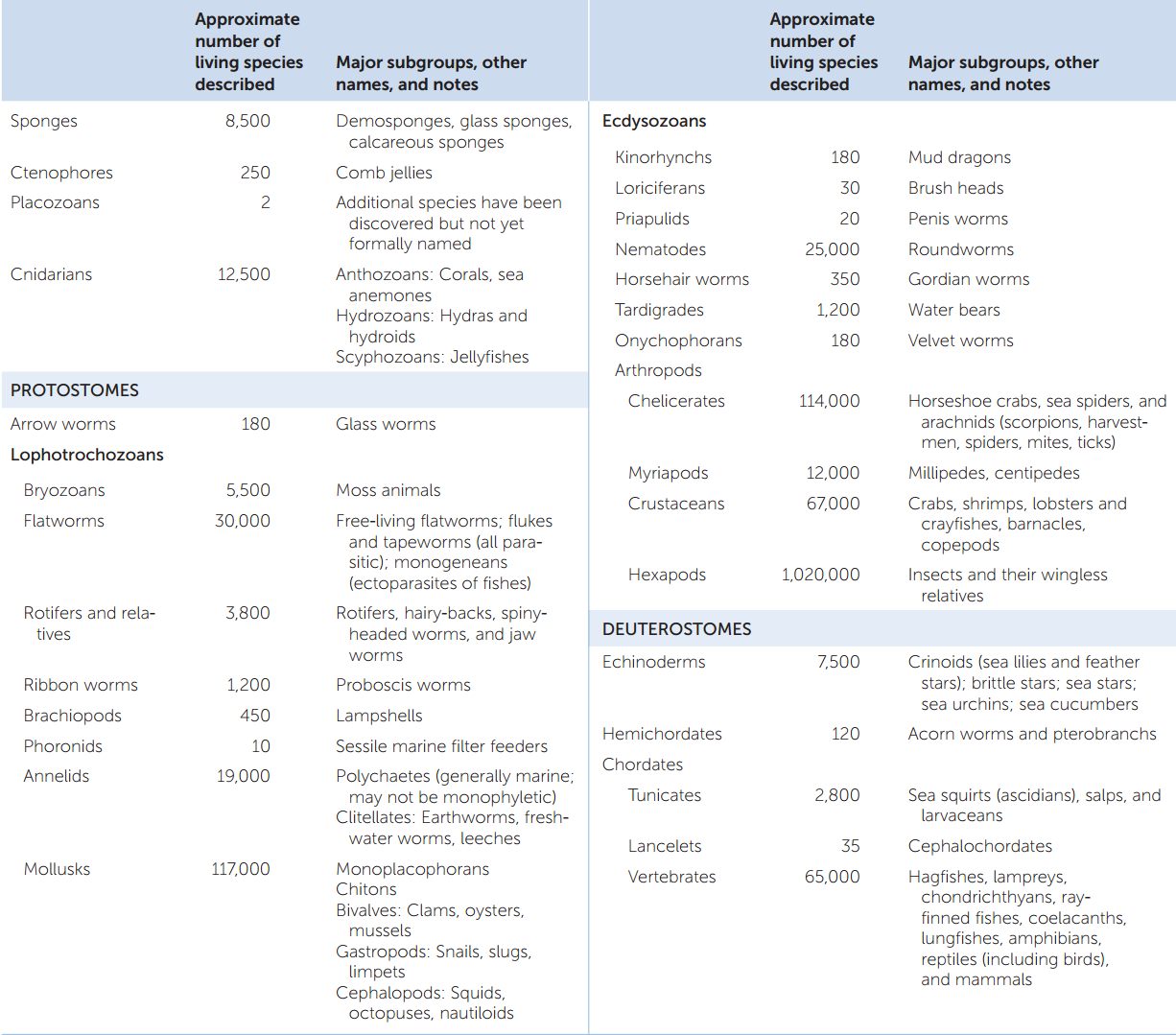
The most convincing evidence that all the organisms considered to be animals share a common ancestor comes from phylogenetic analyses of their gene sequences. Relatively few complete animal genomes are available, but more are being sequenced each year. Analyses of these genomes and of many individual gene sequences have shown that all animals are indeed monophyletic. A currently well supported phylogenetic tree of the animals is shown in FIGURE 23.1.
Surprisingly few morphological features are shared across all animal species. The morphological synapomorphies (shared, derived traits) that are present are evident primarily at the cellular level. They include several unique features of the junctions between animal cells and a common set of extracellular matrix molecules, including collagen and proteoglycans. Although some animals in a few groups lack one or another of these cellular characteristics, it is believed that these traits were possessed by the ancestor of all animals and were subsequently lost in those groups.
471
LINK
Animal cell junctions and extracellular matrix molecules are described in Concept 4.5
Similarities in the organization and function of Hox and other developmental genes provide additional evidence of developmental mechanisms shared by a common animal ancestor. The Hox genes specify body pattern and axis formation, leading to developmental similarities across animals (see Chapter 14).
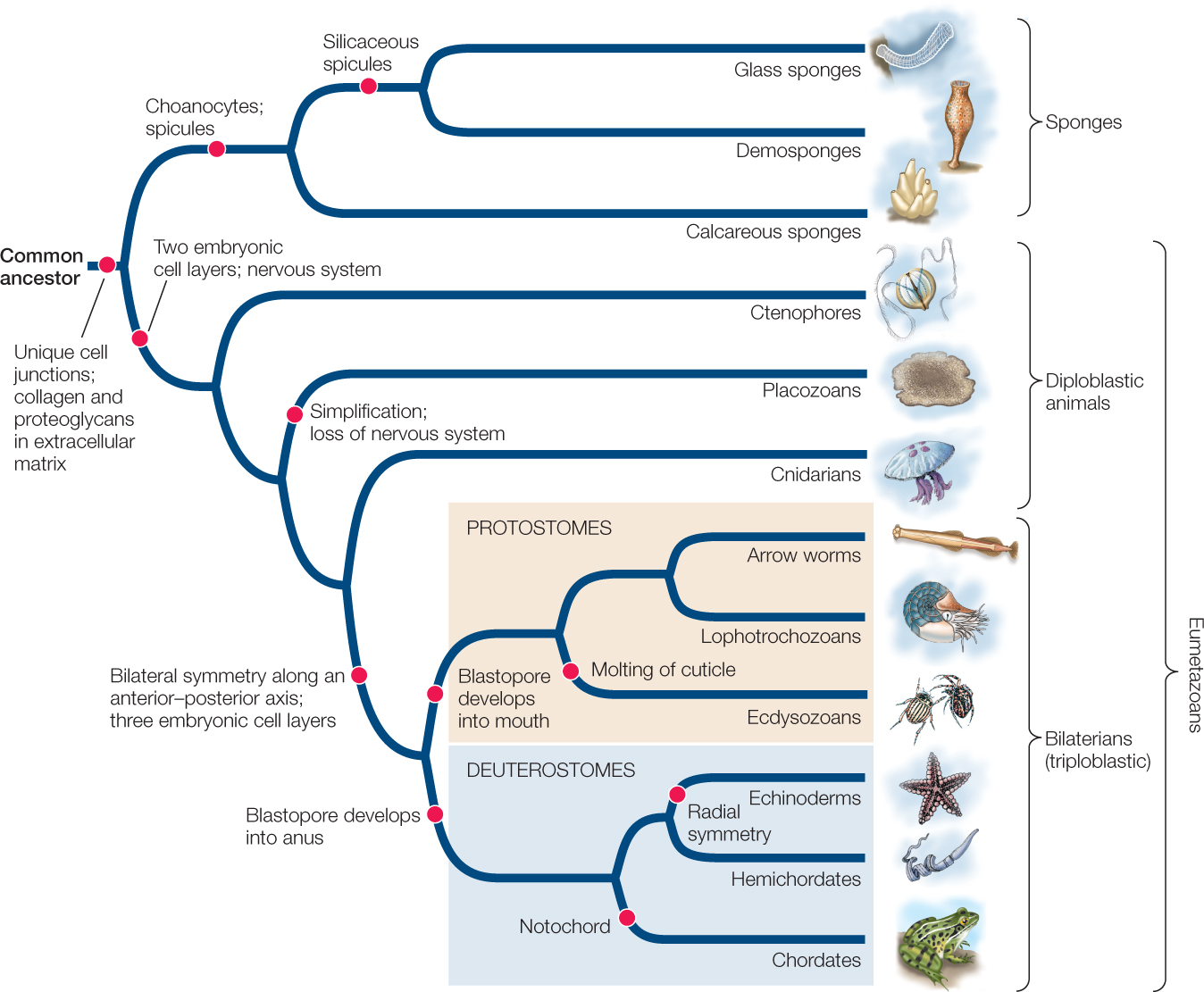
The common ancestor of animals was probably a colonial flagellated protist similar to existing colonial choanoflagellates (FIGURE 23.2A), which have similarities to the multicellular sponges (FIGURE 23.2B). Why did these early animal ancestors begin to form multicellular colonies? One hypothesis is that multicellular colonies are more efficient than are single cells at capturing their prey. Experiments with living species of choanoflagellate show that they spontaneously form multicellular colonies in response to signaling compounds that are found on certain species of planktonic bacteria they eat (FIGURE 23.3).
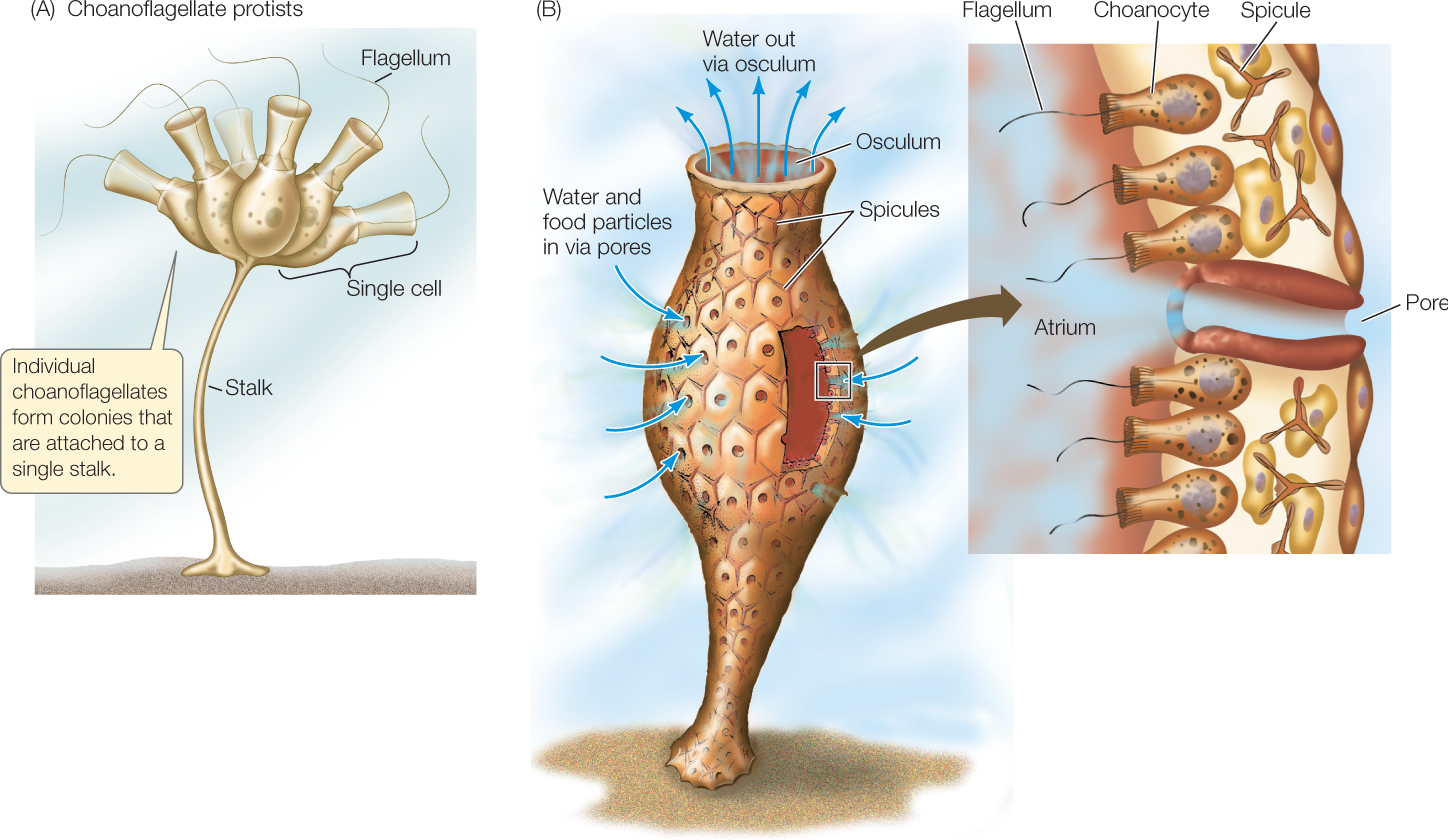
Investigation
HYPOTHESIS
Multicellular colonies of S. rosetta are induced by chemical compounds found on specific species of planktonic bacteria.
- Isolate individual species of bacteria from an environmental sample of S. rosetta and its associated bacterial community.
- Use a combination of antibiotics to produce a bacteria-reduced culture line of S. rosetta that does not spontaneously form rosette colonies (the RCA line, for “rosette colonies absent”).
- Feed the RCA line with each of the isolated species of bacteria from step 1. Record any rosette colony formation.
- Isolate compounds that induce multicellular colony formation from rosette-producing bacteria identified in step 3.
Alegado and colleagues tested various compounds from Algoriphagus and found that RIF-1, a sulfonolipid produced by the bacteria, is the signal that induces formation of multicellular colonies in S. rosetta.
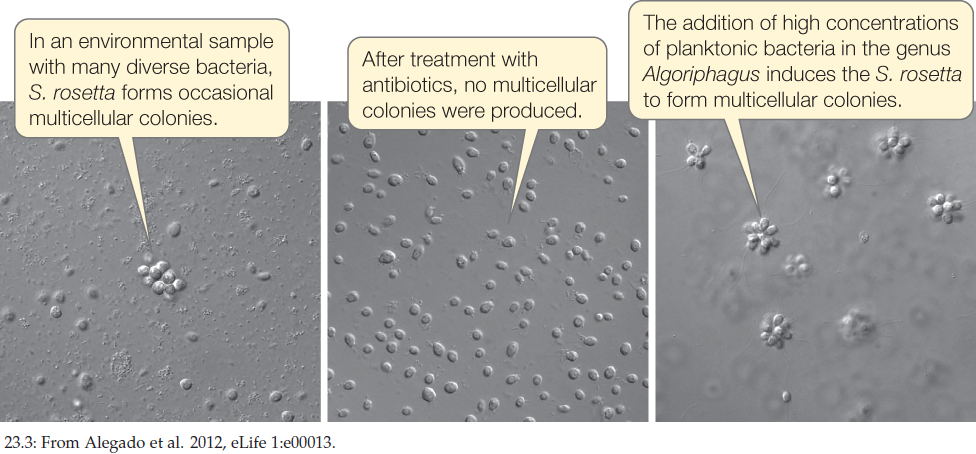
CONCLUSION
A chemical signal (RIF-1) from a particular species of prey bacteria can induce the formation of multicellular colonies in some species of choanoflagellates.
aR. A. Alegado et al. 2012. eLife 1:e00013.
Once multicellular colonies were formed in the ancestral animal lineage, it is likely that certain cells in the colony began to be specialized—some for movement, others for nutrition, others for reproduction, and so on. Once this functional specialization had begun, cells could continue to differentiate. Coordination among groups of cells could have improved by means of specific regulatory and signaling molecules that guided differentiation and migration of cells in developing embryos. Such coordinated groups of cells eventually evolved into the larger and more complex organisms that we call animals.
Nearly 80 percent of the 1.8 million named species of living organisms are animals, and millions of additional animal species await discovery. Evidence about the evolutionary relationships among animal groups can be found in fossils, in patterns of embryonic development, in the morphology and physiology of living animals, in the structure of animal proteins, and in gene sequences. Increasingly, studies of higher-level relationships have come to depend on genomic sequence comparisons, as genomes are ultimately the source of all inherited trait information.
Basic developmental patterns and body plans differentiate major animal groups
Distinct layers of cells form during the early development of most animals. These cell layers differentiate into specific organs and organ systems as development continues. The embryos of diploblastic animals have two cell layers: an outer ectoderm and an inner endoderm. Embryos of triploblastic animals have, in addition to ectoderm and endoderm, a third distinct cell layer, mesoderm, between the ectoderm and the endoderm. The existence of three cell layers in embryos is a synapomorphy of triploblastic animals, whereas the diploblastic animals (ctenophores, placozoans, and cnidarians) retain the ancestral condition. Some biologists consider sponges to be diploblastic, but since they do not have clearly differentiated tissue types or embryonic cell layers, the term is not usually applied to them.
472
During early development in many animals, in a process known as gastrulation, a hollow ball one cell thick indents to form a cup-shaped structure. The opening of the cavity formed by this indentation is called the blastopore:
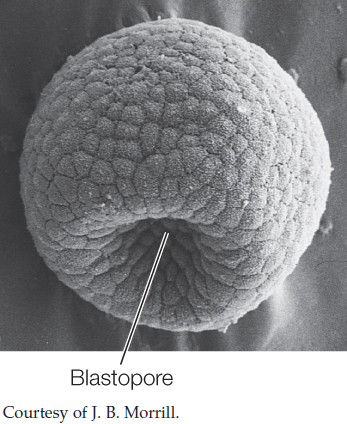
Gastrulation will be covered in detail in Chapter 38. The point to remember here is that the overall pattern of gastrulation immediately after the blastopore stage divides the triploblastic animals into two major groups:
- In the protostomes (Greek, “mouth first”), the mouth arises from the blastopore, and the anus forms later.
- In the deuterostomes (“mouth second”), the blastopore becomes the anus, and the mouth forms later.
Although the developmental patterns of animals are more varied than suggested by this simple dichotomy, phylogenies based on DNA sequences indicate that the protostomes and deuterostomes are distinct clades. These groups are known as the bilaterians (named for their usual bilateral symmetry), and they account for the vast majority of animal species.
The general structure of an animal, the arrangement of its organ systems, and the integrated functioning of its parts are referred to as its body plan. As Chapter 14 described, the regulatory and signaling genes that govern the development of body symmetry, body cavities, segmentation, and appendages are widely shared among the different animal groups. Although the myriad animal body plans cover a wide range of morphologies, they can be seen as variations on four key features:
- The symmetry of the body
- The structure of the body cavity
- The segmentation of the body
- The existence and location of external appendages that are used for sensing, feeding, locomotion, mating, and other functions
Each of these features affects how an animal interacts with its environment.
Most animals are symmetrical
The overall shape of an animal can be described by its symmetry. An animal is said to be symmetrical if it can be divided along at least one plane into similar halves. Animals that have no plane of symmetry are asymmetrical. Placozoans and many sponges are asymmetrical, but most other animals have some kind of symmetry, which is governed by the expression of regulatory genes during development.
In organisms with radial symmetry, body parts are arranged around a single axis at the body’s center:
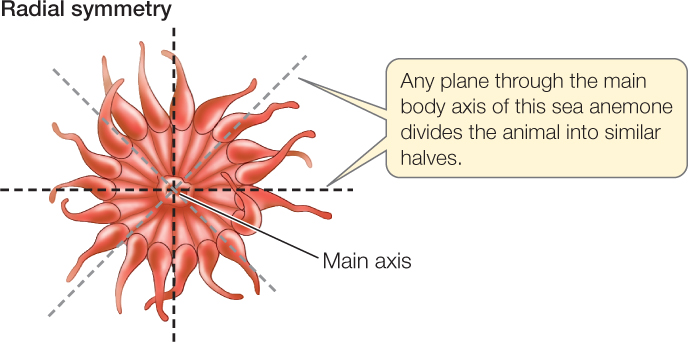
A perfectly radially symmetrical animal can be divided into similar halves by any plane that contains the central axis. However, most radially symmetrical animals are slightly modified, so fewer planes can divide them into identical halves. Some radially symmetrical animals are sessile (they remain fixed in one place) or drift with water currents. Others move slowly but can move equally well in any direction.
Bilateral symmetry is characteristic of animals that have a distinct front end, which typically precedes the rest of the body as the animal moves. A bilaterally symmetrical animal can be divided into mirror-image (left and right) halves by a single plane that passes through the midline of its body. This plane runs from the front, or anterior, end of the body, to the rear, or posterior, end:
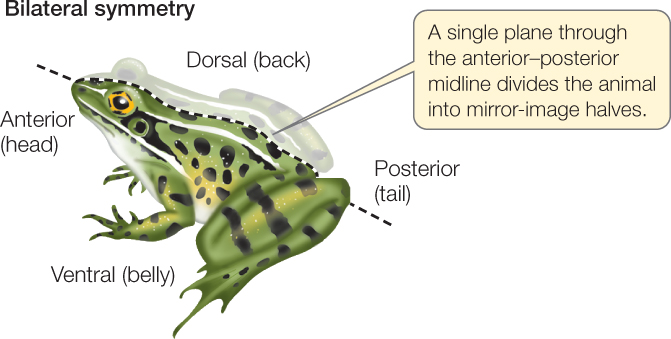
A plane at right angles to the midline divides the body into two dissimilar sides, its dorsal (top) and ventral (bottom) halves.
Bilateral symmetry is strongly correlated with cephalization (Greek kephalos, “head”), which is the concentration of sensory organs and nervous tissues at the anterior end of the animal. Cephalization has been evolutionarily favored because the anterior end of a bilaterally symmetrical animal typically encounters the environment first.
The structure of the body cavity influences movement
Animals with three embryonic cell layers (see Figure 23.1) can be divided into three types based on the presence and structure of an internal, fluid-filled body cavity.
- Acoelomate animals such as flatworms lack an enclosed, fluid-filled body cavity. Instead, the space between the gut (derived from endoderm) and the muscular body wall (derived from mesoderm) is filled with masses of cells called mesenchyme (FIGURE 23.4A). These animals typically move by beating cilia.
- Pseudocoelomate animals have a body cavity called a pseudocoel, a fluid-filled space in which many of the internal organs are suspended. A pseudocoel is enclosed by muscles (mesoderm) only on its outside; there is no inner layer of mesoderm surrounding the internal organs (FIGURE 23.4B).
- Coelomate animals have a body cavity, the coelom, that develops within the mesoderm. It is lined with a layer of muscular tissue called the peritoneum, which also surrounds the internal organs. The coelom is thus enclosed on both the inside and the outside by mesoderm (FIGURE 23.4C).
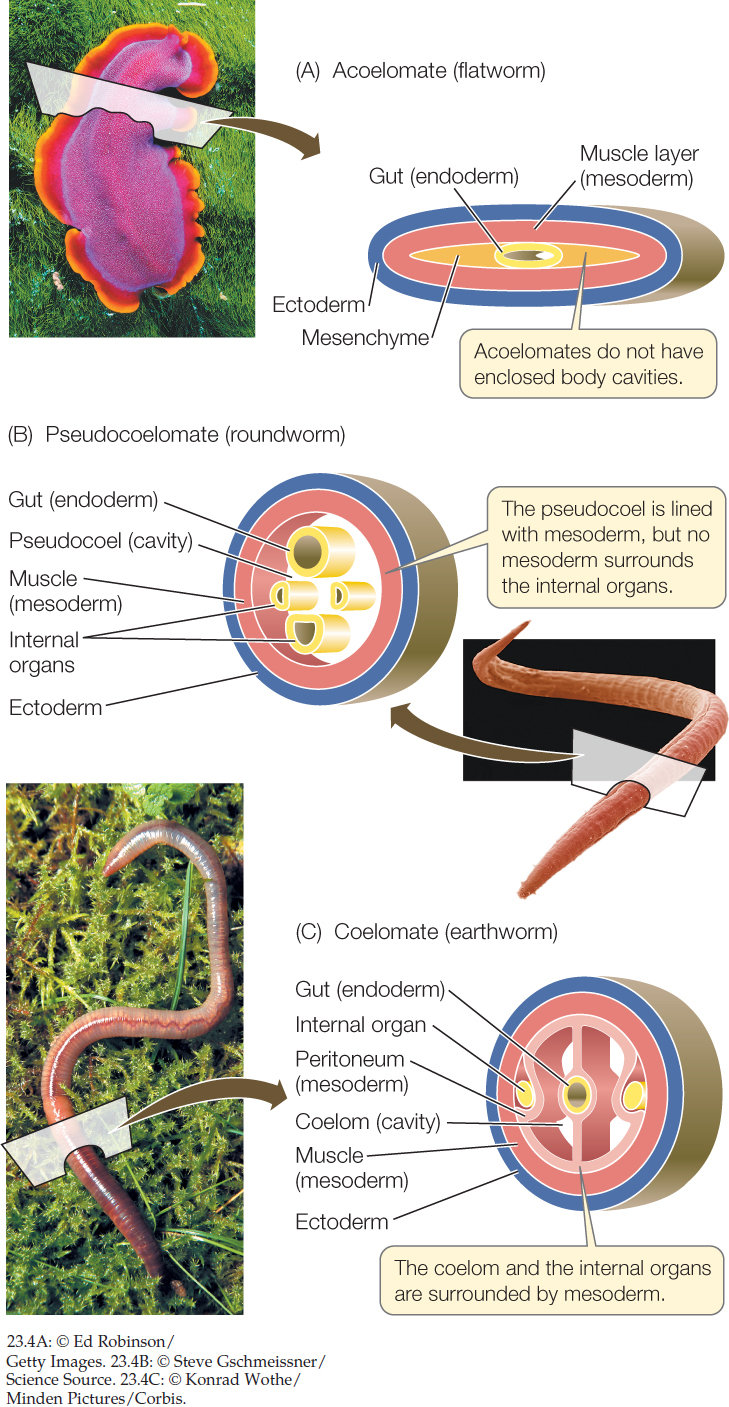
The structure of an animal’s body cavity strongly influences the ways in which it can move. The body cavities of many animals function as hydrostatic skeletons. Fluids are relatively incompressible, so when the muscles surrounding a fluid-filled body cavity contract, fluids shift to another part of the cavity. If the body tissues around the cavity are flexible, fluids squeezed out of one region can cause another region to expand. The moving fluids can thus move specific body parts. (You can see how a hydrostatic skeleton works by watching a snail emerge from its shell.) A coelomate animal has better control over the movement of the fluids in its body cavity than a pseudocoelomate animal does. An animal that has longitudinal muscles (running along the length of the body) as well as circular muscles (encircling the body cavity) has even greater control over its movement.
In terrestrial environments, the hydrostatic function of fluid-filled body cavities applies mostly to relatively small, soft-bodied organisms. Most larger animals (as well as many smaller ones) have hard skeletons that provide protection and facilitate movement. Muscles are attached to those firm structures, which may be inside the animal or on its outer surface (in the form of a shell or cuticle).
Segmentation improves control of movement
Segmentation—the division of the body into segments—is seen in many animal groups. Segmentation facilitates specialization of different body regions. It also allows an animal to alter the shape of its body in complex ways and to control its movements precisely. If an animal’s body is segmented, muscles in each individual segment can change the shape of that segment independently of the others. In only a few segmented animals is the body cavity separated into discrete compartments, but even partly separated compartments allow better control of movement. Segmentation occurs in several groups of protostomes and deuterostomes.
474
In some animals, such as annelids (earthworms and their relatives), similar body segments are repeated many times (FIGURE 23.5A). In other animals, including most arthropods, segments differ strikingly from one another (FIGURE 23.5B). The dramatic evolutionary radiation of the arthropods (including the insects, spiders, centipedes, and crustaceans) was based on changes in a segmented body plan that features muscles attached to the inner surface of an external skeleton, including a variety of external appendages that move these animals. In some animals, distinct body segments are not apparent externally—for example, the segmented vertebrae of vertebrates, including humans. Nonetheless, muscular segmentation is clearly visible in humans with well-defined, muscular bodies (FIGURE 23.5C).

Appendages have many uses
Getting around under their own power allows animals to obtain food, to avoid predators, and to find mates. Even some sedentary species, such as sea anemones, have larval stages that use cilia to swim, thus increasing the animal’s chances of finding a suitable habitat.
Appendages that project externally from the body greatly enhance an animal’s ability to move around. Many echinoderms, including sea urchins and seas stars, have large numbers of tube feet that allow them to move slowly across the substrate. Animals whose appendages have become modified into limbs are capable of highly controlled, more rapid movement. The presence of jointed limbs has been a prominent factor in the evolutionary success of the arthropods and the vertebrates. In four independent instances—among the arthropod insects and among the vertebrate pterosaurs, birds, and bats—body plans emerged in which limbs were modified into wings, allowing these animals to use powered flight.
Appendages also include many structures that are not used for locomotion. Many animals have antennae, which are specialized appendages used for sensing the environment. Other appendages (such as claws and the mouthparts of many arthropods) are adaptations for capturing prey or chewing food. In some species, appendages are used for reproductive purposes, such as sperm transfer or egg incubation.
Nervous systems coordinate movement and allow sensory processing
The bilaterian animals have a well-coordinated central nervous system. More diffuse nervous systems, called nerve nets, are present in some other animals, such as jellyfishes and their relatives. Nervous systems appear to be completely absent in a few sessile animal groups, such as sponges.
The central nervous system of bilaterians coordinates the actions of muscles, which allows coordinated movement of appendages and body parts. This coordination of muscles permits highly effective and efficient movement on land, in water, or through the air. The central nervous system is also essential for the processing of sensory information gathered from a wide variety of sensory systems. Many animals have sensory systems for detecting light, for forming images of their environment (sight), for mechanical touch, for detecting movement, for detecting sounds (hearing), for detecting electrical fields, and for chemical detection (e.g., taste and smell). These sensory systems allow animals to find food, and the ability of animals to move allows them to capture or collect food from their environment. These same abilities also allow most animals to move to avoid potential predators or to search for suitable mates. Most animals can also assess the suitability of different environments and move appropriately in response to that information.
475
CHECKpointCONCEPT23.1
- Describe the difference between diploblastic and triploblastic embryos, and between protostomes and deuterostomes.
- Describe the main types of symmetry found in animals. How can an animal’s symmetry influence the way it moves?
- Explain several ways in which body cavities and segmentation improve control over movement.
Variations in body symmetry, body cavity structure, life cycles, patterns of development, and survival strategies differentiate the many animal species. TABLE 23.1 summarizes the living members of the major animal groups we will describe in the rest of this chapter.