CONCEPT38.6 Development Continues throughout Life
No sexually produced animal is as large as its parent at birth or hatching. All must obtain nourishment and grow. Some animals immediately look like their parents and undergo direct development to reach full size. Others appear very different and spend part of their lives as one or more different forms, as they undergo indirect development.
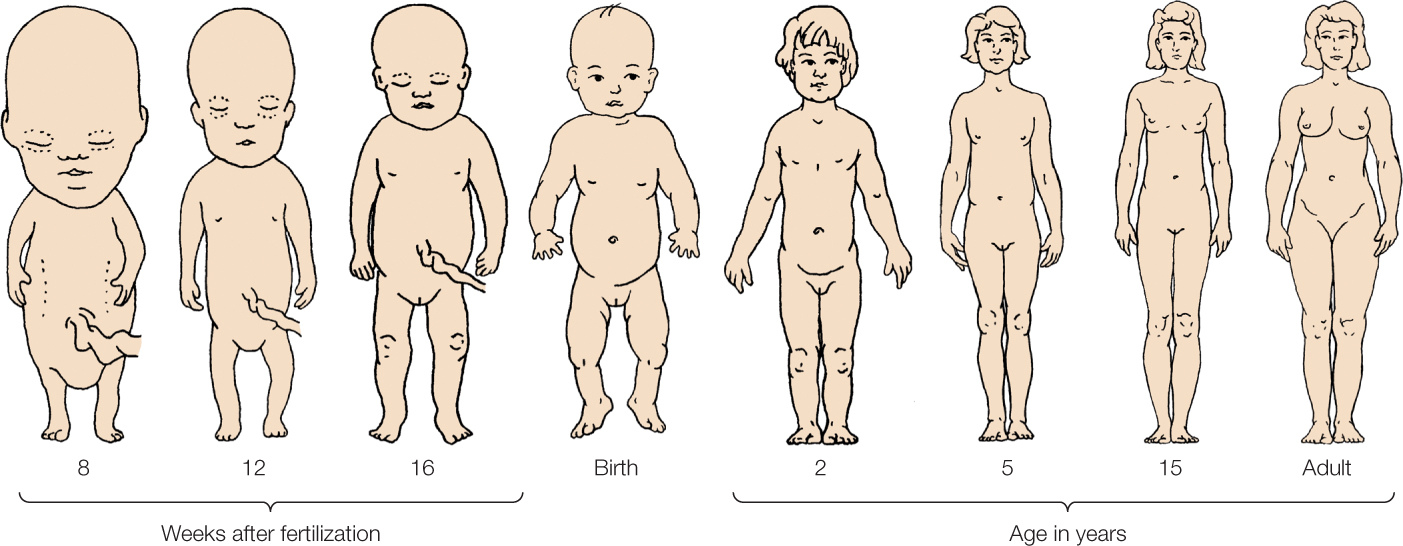
A change in the proportions of various parts of an organism relative to one another as a consequence of unequal growth is known as allometry. Isometric growth is a 1:1 size increase; allometric growth is any change from this. Allometry is often seen in direct developers as parts of their body change in relative size and shape with growth. For example, at birth a human baby is not a miniature adult. Not only are many of its systems immature, but its overall morphology (form and shape) is also quite different than that of an adult. One difference is the ratio of head to trunk size. A baby’s head is much larger in comparison with the rest of its body than an adult’s head is, and the head stops growing much earlier in life than the rest of the body (FIGURE 38.21). It has been suggested that the large head of infant mammals, especially humans, corresponds to the large brain and development of the cerebral cortex. Many examples of allometry exist. Indeed, nearly all direct developers experience allometric growth of some sort.
Indirect development involves changes in morphology that can be dramatic. Many animals, both protostome and deuterostome, have a larval stage that may be quite unlike the adult. A familiar example is the monarch butterfly, which has a caterpillar that feeds on the milkweed plant on which its egg was placed, and a winged adult that can migrate from Canada to Mexico. FIGURE 38.22 shows two larvae with which you might not be familiar.
805

A larval stage usually performs one of two important roles—or both. Like the caterpillar, it may be primarily a feeding stage. This is seen in animals that provision their eggs with little or moderate yolk resources. Insects, fish, and amphibians are prime examples of this lifestyle. Some adult insects do not feed at all. Ephemeridae—mayflies—last only 1 day as adults. (They are ephemeral, as their name tells us.) They molt from the pupal to adult stage, seek a mate, deposit eggs, and die, relying solely on energy stored by the larva.
Larvae may also play an important role in species distribution. This is especially true in animals in which the adult is sedentary or relatively so. Corals, for example, can reproduce by fragmentation, but new sites are colonized by swimming larvae. Some polychaete worms (capitellids) are rapid colonizers of freshly disturbed, organically rich areas (substrates of polluted harbors and beneath fish farms, oil spills, and whale falls—decaying whale carcasses on the ocean floor) that are widely distributed around the world. Larvae, swimming only with ciliary bands, are carried by currents and then settle in these areas.
When a larva transforms into an adult at metamorphosis, changes can be dramatic, as in the monarch and other butter-flies and moths. Major portions of the body may be remodeled or even discarded (see Concept 23.4). Many amphibians lay their eggs in spring, and after quite rapid development, small tadpoles emerge. A tadpole feeds, and its size increases. At metamorphosis, the tadpole shows changes in virtually every system. Tail and gills are resorbed, and limbs grow. In many species, the tadpole stage is herbivorous whereas the adult is carnivorous. Accompanying this change is an alteration of gut length. The gut of a tadpole is actually several times longer than the gut of the small froglet it will become! A tunicate (sea squirt; see Figure 23.35A) tadpole settles and rapidly resorbs its tail, including the notochord and nerve cord. Since the three major chordate characteristics (notochord, hollow dorsal nerve chord, and post-anal tail) are no longer present, the adult tunicate does not resembles a chordate! The larva of the lamprey is so different from the adult that it was once considered a different species. It is still called by the earlier name of “ammocoetes.”
CHECKpointCONCEPT38.6
- Prior to the Renaissance, painters often depicted infants as miniature adults. Why do these babies not appear natural?
- In addition to head size, give another example of allometric growth in humans.
- What are two major functions of larvae?
As we have seen, the wonderful process that we recognize as development begins with the fusion of a single egg and a single sperm and continues throughout the entire life of the organism. Although not considered here, the processes of regeneration, wound healing, and aging are all developmental mechanisms. Medical interests in stem cell technology, in wound repair and regeneration, in the prevention of birth defects, and in many other areas, as well as a strong interest in the overlapping field of evolutionary biology, are adding impetus to our desire to understand developmental processes in many organisms. With all of the exciting techniques now available, the door for future studies is wide open.
806
How does the Sonic hedgehog pathway control development of the vertebrate brain and eyes?
ANSWER Sonic hedgehog (Shh) is a transcription factor secreted by the notochord (Concept 38.4). It diffuses to the overlying neural tube, where it induces differentiation of the ventral neural tube into motor neurons and other features of the ventral regions of the spinal cord. Shh also plays different inductive roles at other locations. At the anterior end of the neural tube where the forebrain will develop, the closed neural tube undergoes reciprocal inductive tissue interactions with the overlying ectoderm. Signaling molecules from the neural tissue induce the ectoderm to form the lens of the eye, and the developing lens produces signaling molecules that induce the underlying neural tissue to form an optic cup that will differentiate into the retina.
But why do these events happen bilaterally? Mesoderm that lies anterior to the notochord produces and releases Shh, as does the notochord. Shh induces the ventral region of the anterior neural tube to differentiate into the structures of the forebrain. One of the actions of Shh is to inhibit a transcription factor called Pax6 that is expressed by the neural tissue. Pax6 is essential for determination of the eye field (i.e., tissues that can form eyes; see Figure 14.22). The midline expression of Shh splits the eye field into two and also enables the forebrain to develop into left and right hemispheres.
When environmental factors affect the action of Shh, holoprosencephaly can result. When pregnant cows or sheep graze on Veratrum californicum at certain critical points in gestation, the embryo is exposed to cyclopamine and the Shh signaling pathway is blocked. Depending on when the exposure occurs, bilateral development of the forebrain may not take place, resulting in holoprosencephaly and cyclopia (FIGURE 38.23). Another area of great interest for humans is the observation that Shh requires cholesterol to be an effective signal. Abnormally low levels of cholesterol during pregnancy (such as might occur with the use of certain statin drugs) could cause some aspects of holoprosenecephaly.
