CONCEPT41.3 Biogeography Reflects Physical Geography
An organism’s physiology, morphology, and behavior—all aspects of its phenotype—affect how well it can tolerate a particular physical environment. For this reason, the physical environment of a place greatly influences what species can live there. Furthermore, we expect species that occur in similar environments to have evolved similar phenotypic adaptations to those conditions.
Similarities in terrestrial vegetation led to the biome concept
The European scientist–explorers who traveled the globe noticed that the vegetation found in climatically similar regions on different continents was strikingly convergent. This observation led to the concept of a biome: a distinct physical environment that is inhabited by ecologically similar organisms with similar adaptations. The species that occupy the same biome in geographically separate regions are often not closely related. Their similarities therefore reflect convergent evolution by natural selection (see Concept 16.1).
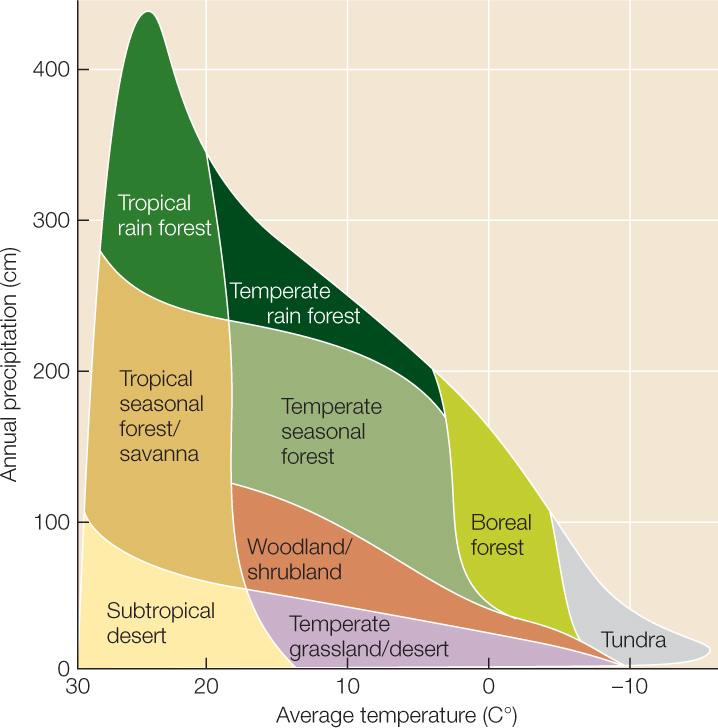
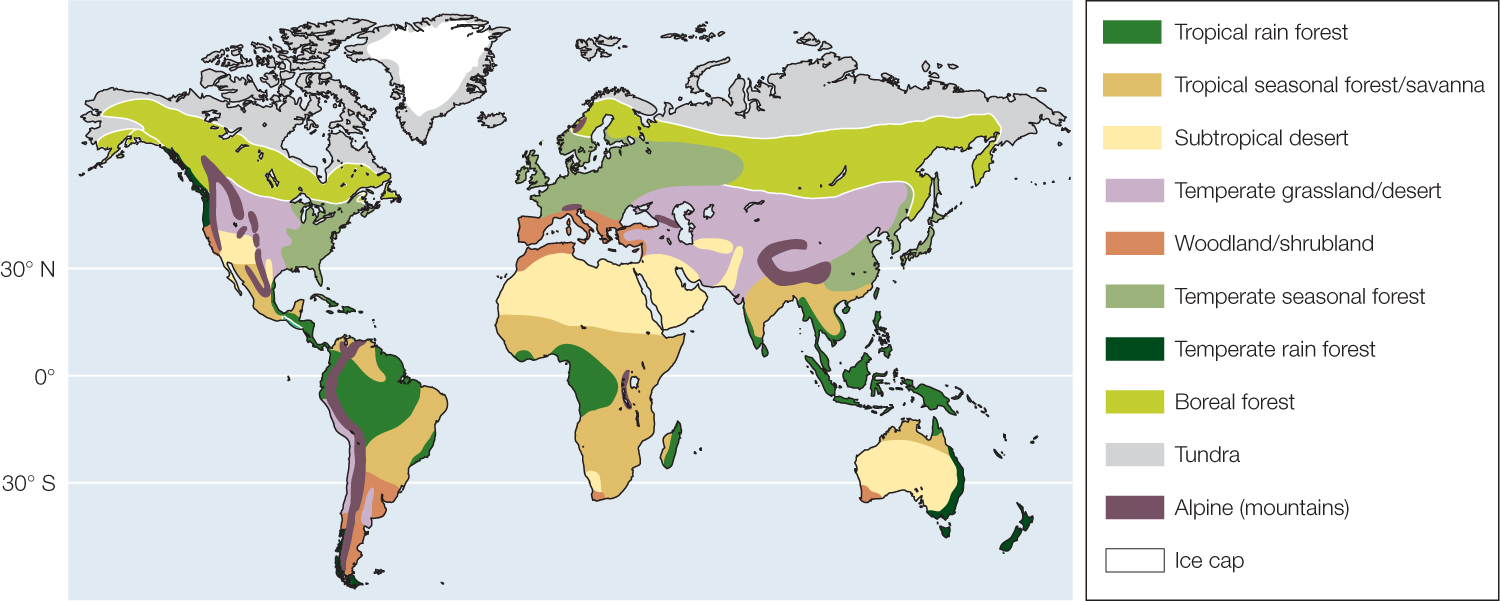
Terrestrial biomes are generally distinguished by the characteristics of their vegetation—whether the dominant plants are woody trees or shrubs, grasses, or broad-leaved herbaceous plants, and whether their leaves are big or small, deciduous (dropped seasonally) or evergreen. Characteristics of organisms other than plants also distinguish the biomes, since these characteristics represent adaptations to the vegetation as well as the climate. The distribution of terrestrial biomes is broadly determined by annual patterns of temperature and precipitation (FIGURE 41.10). Latitudinal gradients in average temperature and differences in its seasonal fluctuation separate tropical, subtropical, temperate, boreal, and polar biomes. Within those broad temperature zones, average annual precipitation and its pattern of seasonal fluctuation predict what biome will be found. In tropical biomes, for example, the length of the dry season determines whether tropical rainforest, tropical seasonal forest, or savanna develops. Elevation-associated gradients in temperature also determine the distribution of biomes; for example, tundra can be found on mountaintops, even in the tropics (FIGURE 41.11).
LINK
Some plant species can tolerate the cold temperatures of the tundra. You can find out more about plant adaptations to various environments in Concept 28.3
Climate is not the only factor that molds terrestrial biomes
Other features of the physical environment interact with climate to influence the character of terrestrial vegetation. Southwestern Australia, for example, has a “Mediterranean” climate with hot, dry summers and cool, moist winters. This climate generally supports woodland/shrubland vegetation. Succulent plants, which are well adapted to summer drought, are common in Mediterranean climates—but not in Australia, which has nutrient-poor soils. The lack of nitrogen in these soils makes it expensive for plants to construct the nitrogen-rich photosynthetic machinery of leaves. As a result, many plants produce long-lived leaves that are defended against herbivores in ways that do not require nitrogen (see Concept 28.2). These defenses include tough, indigestible cell walls, compounds that reduce the nutritional quality of leaves, and toxic oils and resins—features that also make the vegetation highly flammable. Plant material produced during the winter rains dries out during the summer and fuels intense fires that periodically sweep across the landscape. As a result, succulent plants, which are easily killed by fires, are rare in southwestern Australia.
855
Soil nutrients and fire bring us back to the desert grasslands of the U.S.–Mexico Borderlands. Grasslands—areas dominated by grasses and herbs rather than trees or shrubs—normally occur where precipitation is too scanty, or too sporadic, to support forests but is more plentiful than is typical of deserts. Fully one-fourth of Earth’s land surface experiences such conditions, and much of this area supports grasslands. But grasslands are also found in some places where we do not expect them (FIGURE 41.12). Their presence in these places demonstrates that biome boundaries are fuzzy and are not perfectly predicted by temperature and precipitation because other factors also affect the character of vegetation.

856
One such factor is fire, which rarely kills grasses but often kills shrubs and trees. Before humans arrived, the U.S.–Mexico Borderlands experienced periodic natural wildfires, ignited by lightning, that produced desert grassland rather than desert shrubland (see Figure 41.12A). Humans themselves have contributed to the establishment of grasslands in many areas. Fires lit by humans in the Serengeti Plain of northern Tanzania, Africa, may have helped convert the landscape from tropical seasonal forest to savanna, a biome in which grasses dominate and trees are widely scattered (see Figure 41.12B). Such human effects are not necessarily recent. Humans have inhabited Australia for at least 50,000 years, for instance, and their fires appear to have transformed some of the original woodlands in the arid “red center” of the continent into grasslands dominated by “spinifex” grasses (genus Triodia; see Figure 41.12C). Ecosystems in which grasses dominate are also found in other places where we might expect to find forests. When humans brought domesticated livestock into temperate regions such as the European Alps, they often converted forests into grassy meadows to feed their animals (see Figure 41.12D).
The properties of soils interact with climate, fire, or grazing to affect the character of vegetation. Trees and shrubs grow particularly slowly in nutrient-poor soils. Lack of sufficient nutrients might prevent a tree or shrub seedling from sending its roots deep enough in the soil to survive the next dry season, or from growing large enough to survive the next wildfire, or from being able to recover from being eaten. If so, then the availability of soil nutrients might tip the balance between woody vegetation and grassland in seasonally dry climates, fire-prone regions, or grazing lands. The examples of grasslands we have chosen (see Figure 41.12) do differ in their soils. Two of them—alpine meadows and the Australian spinifex—grow on relatively nutrient-poor soils, whereas the other two occupy richer volcanic soils.
The biome concept can be extended to aquatic environments
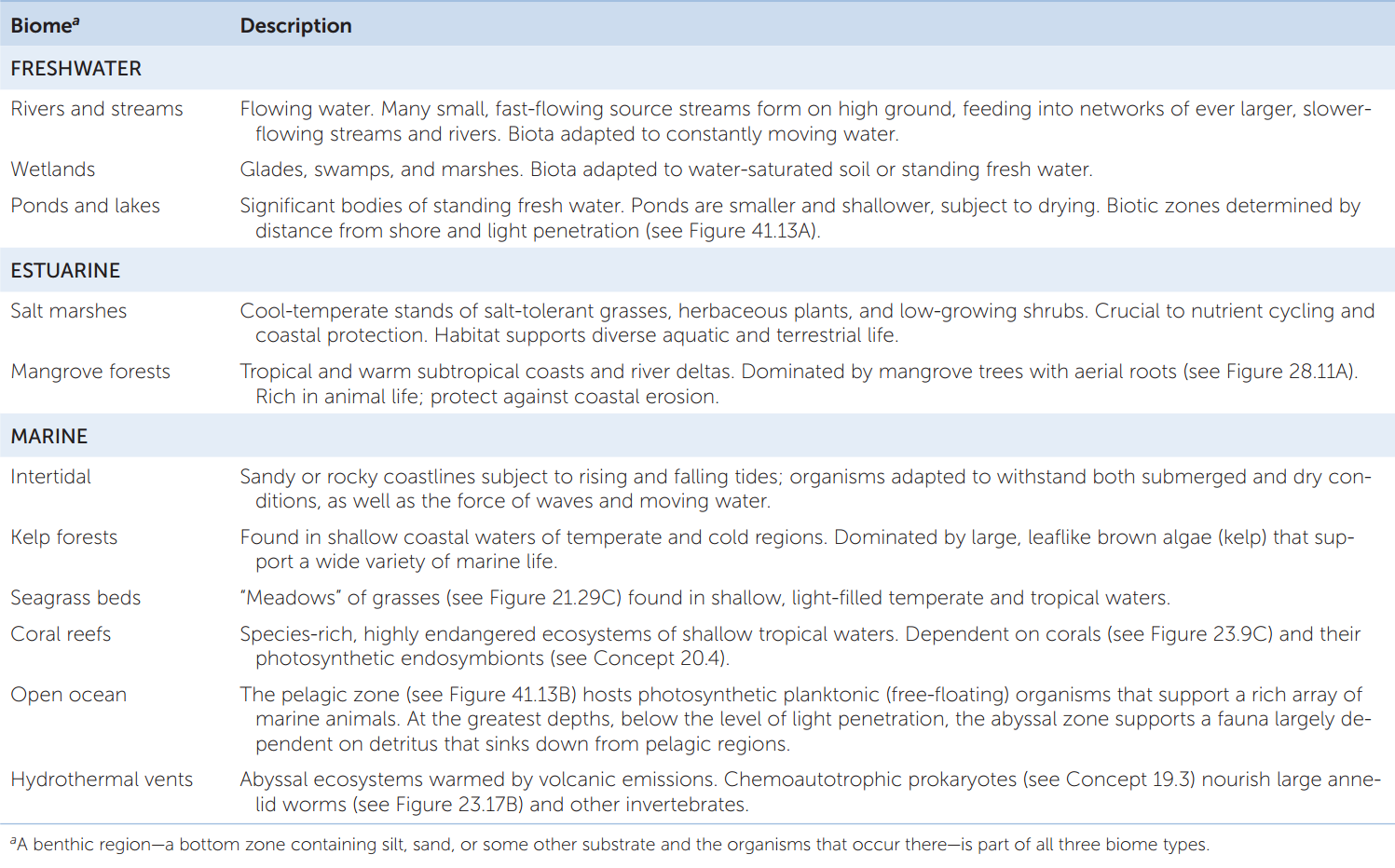
The biome concept can be applied to aquatic environments as well as terrestrial ones (TABLE 41.1), but there are some differences. In aquatic environments, there is no single group of organisms, like terrestrial plants, that is useful for distinguishing biomes. Furthermore, climate (atmospheric conditions of temperature, precipitation, and the like) is less important in determining what organisms grow in a region than are such things as water depth and movement, temperature, pressure, salinity, oxygen content, and the characteristics of the substrate.
857
Salinity distinguishes freshwater biomes (streams, ponds, and lakes), saltwater biomes (salt lakes and oceans), and estuarine biomes (at river mouths where fresh and salt water mix). Salt concentration strongly affects the ability of aquatic organisms to regulate the water content of their bodies (see Concept 36.1) and thus influences which species are found where.
Freshwater biomes can be distinguished by water movement. Streams form wherever precipitation exceeds the amount of water lost to the atmosphere or soil, and the excess water flows downhill. Streams start out small but grow as they join together, forming rivers that eventually reach a lake or ocean in most cases. Within streams, the velocity of the water’s flow determines the strength of the current against which fish must swim and the amount of force that can dislodge bottom-dwelling insects and algae from the streambed. Water flow also determines the presence or absence of sediment. Rapid water flow scours away sediment, exposing rocky surfaces to which organisms can cling. As water slows, it deposits sediment, forming a soft bottom in which organisms can burrow.
Nonflowing biomes such as lakes and oceans can be divided into water-depth zones (FIGURE 41.13). The nearshore regions of lakes (littoral zone) and oceans (littoral or intertidal zone) are shallow, affected by wave action, and periodically exposed to air by fluctuations in water level. Here, different species live in different depth zones according to their tolerance of heat, desiccation, and wave energy.
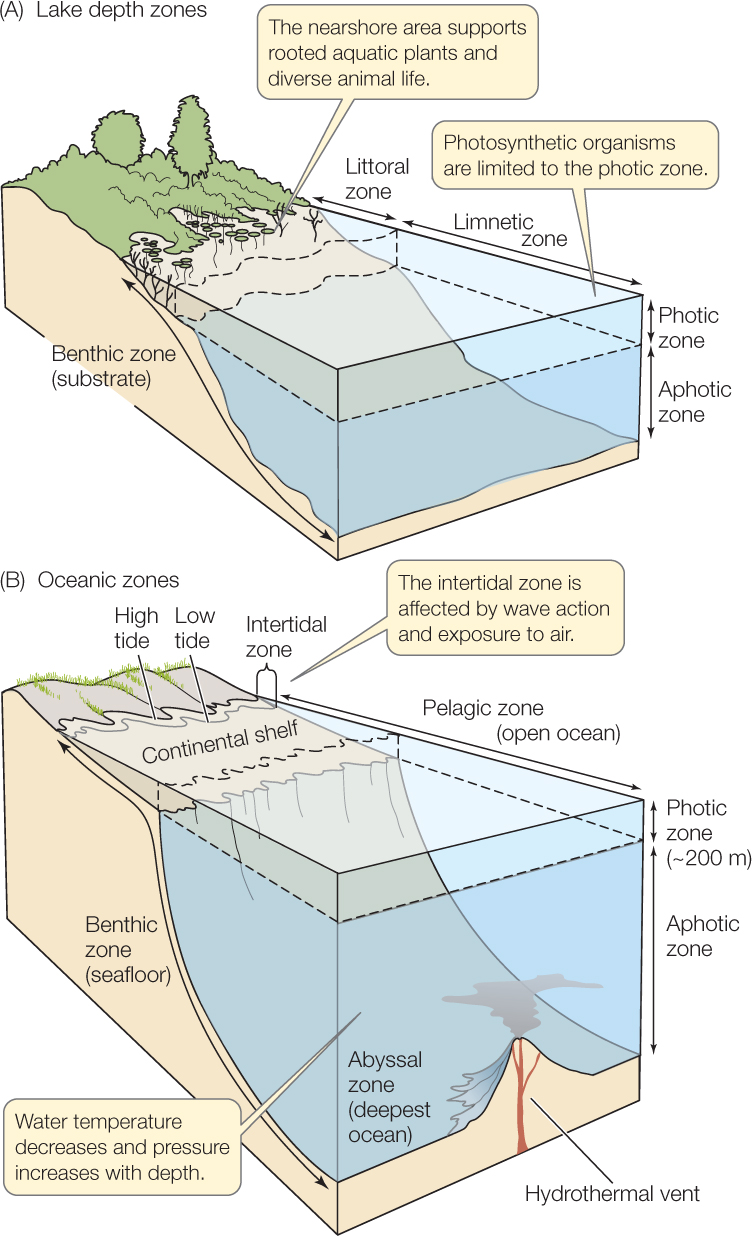
Because only the surface water is in contact with air, dissolved oxygen concentrations are highest in surface waters. Light, too, penetrates only a short distance into water. Photosynthetic organisms are confined to this zone of light penetration, called the photic zone. Rooted aquatic vascular plants and their multicellular algal equivalents (as well as corals with their photosynthetic algal symbionts) grow where the photic zone extends to the bottom, sometimes forming communities with a complex vertical structure much like that of terrestrial plant communities. In the open-water limnetic zone of lakes and the pelagic zone of oceans beyond the continental shelf, the prominent photosynthesizers are phytoplankton (free-floating photosynthetic organisms).
No photosynthetic organisms inhabit the aphotic zone, below the reach of light, which as a consequence is sparsely populated. The lake bottom or ocean floor is called the benthic zone. Water is heavy, so pressure increases with water depth. Temperature generally decreases with depth because cold water is denser than warm water and sinks. Organisms that dwell in the deepest abyssal zone of the oceans experience very high pressures, low oxygen levels, and (except near hydrothermal vents) cold temperatures.
CHECKpointCONCEPT41.3
- What is meant by a biome?
- What conditions limit the distribution of photosynthetic organisms in aquatic biomes? In terrestrial biomes?
- Is it legitimate to consider the human gut a biome? Explain your answer.
We have just seen why physical geography and biogeography are linked. But the distributions of species also are contingent on the movements of continents over Earth’s history.