Eusocial species take social interactions to the extreme
Eusocial A type of social animal in which individuals live in large groups with overlapping generations, cooperation in nest building and brood care, and reproductive dominance by one or a few individuals.
We have seen that many animals are social and interact with conspecifics in a variety of ways. Some animals are so extremely social that we call them eusocial. Eusocial species are distinguished by four characteristics:
- 1. Several adults living together in a group
- 2. Overlapping generations of parents and offspring living together in the same group
- 3. Cooperation in nest building and brood care
- 4. Reproductive dominance by one or a few individuals, and the presence of sterile individuals
Among the insects, eusocial species are limited to the order Hymenoptera, which includes bees, ants, and wasps, and the order Isoptera, which includes the termites (Figure 10.10). Not only are these social insects of evolutionary interest, they are also major players in ecosystem processes. Eusocial insects pollinate plants, consume plant and animal material on a large scale, and recycle wood and organic detritus. Their dominance in the world is due in large part to the immense success of eusociality. Other than insects, the only animals known to be eusocial are two species of mammals that live in underground tunnels in Africa: the naked mole rat (Heterocephalus glaber) and the Damaraland mole rat (Fukomys damarensis).

Caste Individuals within a social group sharing a specialized form of behavior.
Eusocial species are fascinating to ecologists because most individuals in a eusocial group forgo sexual maturation and reproduction. Instead, they specialize at tasks that include defending the group, foraging for the group, or taking care of the subsequent offspring of their parents. Because nonreproductive individuals are specialized for these tasks, they are known as sterile castes. A caste consists of individuals within a social group who share a specialized form of behavior.
How can natural selection produce individuals that lack any reproductive output and therefore lack any direct fitness? To help answer this question, we will start by examining the unique breeding habits of the hymenopteran insects.
Eusociality in Ants, Bees, and Wasps
In Chapter 9 we talked about the unique reproductive behavior of the hymenopteran insects. You may recall that this group of insects makes sons by laying unfertilized eggs and makes daughters by laying fertilized eggs. This method of reproduction is detailed in Figure 10.11. Because one sex is haploid and the other sex is diploid, the hymenopterans are said to have a haplodiploid sex-determination system. As we will see, this sex-determination system helps to favor the evolution of eusociality.
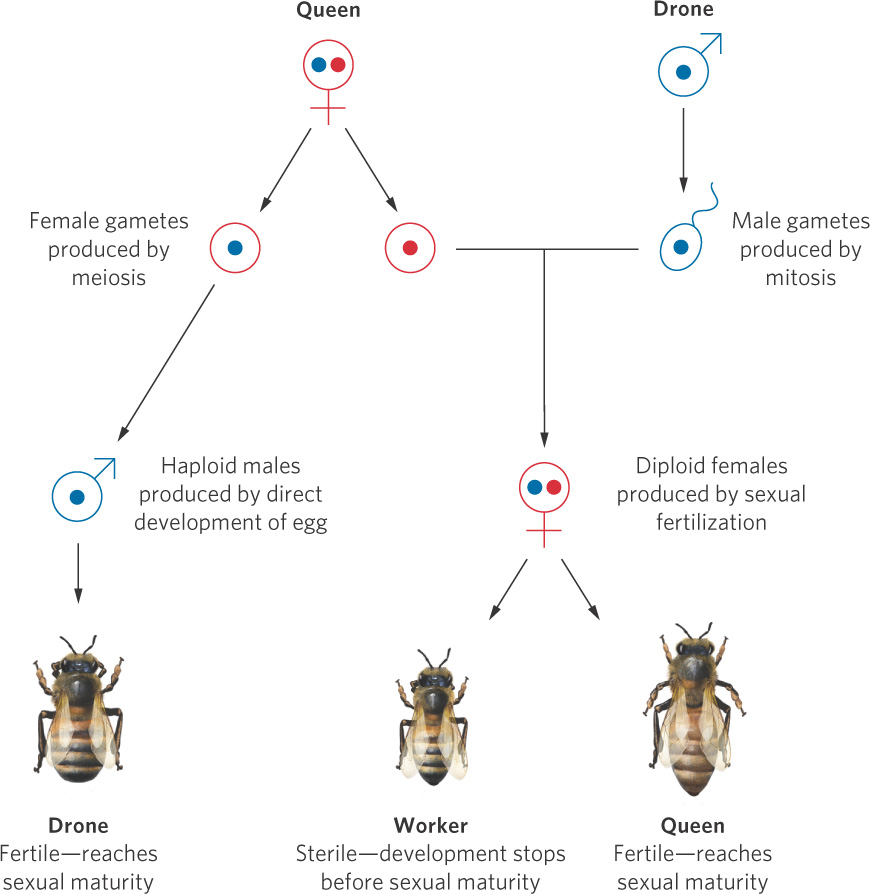
Haplodiploid A sex-determination system in which one sex is haploid and the other sex is diploid.
Social Organization
Queen The dominant, egg-laying female in eusocial insect societies.
Eusocial insect societies are dominated by one egg-laying female or a few egg-laying females, referred to as queens. The queens in colonies of ants, bees, and wasps mate only once during their lives and store enough sperm to produce all their offspring—up to a million or more offspring over a period of 10 to 15 years in some army ants. The nonreproductive progeny of a queen gather food and care for developing brothers and sisters, some of which become sexually mature, leaving the colony to mate and to establish new colonies.
As we discussed in Chapter 9, honeybee societies have a simple organization. The offspring of a queen are divided among a sterile worker caste that is female and a reproductive caste that consists of drones and future queens. Whether an individual becomes a sterile worker or a fertile female is determined by the length of time the larva is fed royal jelly. As a result, the worker caste represents an arrested stage in the development of reproductive females; they stop developing before sexual maturity. Eusocial behavior appears in a large number of hymenopteran species.
241
Coefficients of Relatedness
Haplodiploidy is important to the evolution of eusocial animals because it creates strong asymmetries in coefficients of relatedness. The queen has the same genetic relationship to sons as she has to daughters (r = 0.50), so she can be relatively indifferent to the sex of her reproductive offspring—the drones and new queens that leave the nest—especially when the sex ratio among reproductive individuals in other local hives is nearly equal. However, the relatedness among siblings is quite unique in hymenopterans. If a queen has mated with a single drone, then all females have the same set of genes from their haploid father. This causes females to be more closely related to each other than is typical in a diploid mating system. In the haplodiploid mating system, the coefficient of relatedness between a given female and her sisters is 0.75. In contrast, her brothers are haploid and therefore do not have any genes from the drone, so the average coefficient of relatedness between a given female and her brothers is 0.25. This skewed relatedness favors the evolution of social groups that contain nonreproductive female workers.
To understand how being haplodiploid favors eusociality, we can compare the different options for obtaining fitness in these organisms. For example, a female worker that raises a fertile sister receives the fitness benefit of that individual multiplied by the coefficient of relatedness (r = 0.75). When a female raises its own offspring, it receives the same fitness benefit multiplied by the coefficient of relatedness between a mother and daughter (r = 0.50). This means that the indirect fitness obtained from caring for a sister exceeds the direct fitness that could be obtained from caring for a daughter. Thus, cooperation is likely to be greater among all-female castes than among male castes or, especially, among mixed castes. This may explain why workers in hymenopteran societies are all female.
The fact that workers are three times more closely related to their sisters than to their brothers may also explain why broods of reproductive individuals usually favor females, by about three to one on a weight basis, in spite of the queen’s indifference to the sex ratio. When a female worker can help to rear more female than male reproductive individuals, her own inclusive fitness may actually be higher than it would be if she raised a brood of her own consisting of an equal number of males and females. Under these circumstances, it is not surprising that sterile castes have evolved.
242
Eusociality in Other Species
Although the hymenopterans are among the best known of the eusocial insects, there are a few other species that are also eusocial, including termites and a group of mammals known as naked mole rats. Unlike the haplodiploid hymenopterans, these latter two groups are both diploid.
Termites
Termite colonies can be massive structures that are dominated by a mated pair called the king and queen (Figure 10.12a). Because the queen’s role is almost entirely limited to egg production, her abdomen is grossly distended to allow her to produce thousands of eggs (Figure 10.12b). The king and queen produce sons and daughters by sexual reproduction. With a few exceptions, these sons and daughters can both act as workers in the termite society but they remain sexually immature unless either the king or the queen dies. Many species of termites also have a second caste of nonreproductive individuals known as soldiers. As their name implies, soldiers help to defend the nest against intruders such as ants. Soldiers typically have a very large head that can be used to block the openings of a termite nest to prevent ants from entering.

Mole Rats
There are dozens of species of mole rats, but only two are known to be eusocial: the naked mole rat and the Damaraland mole rat. These rodents live in underground tunnels in African grasslands in colonies of up to 200 individuals. In naked mole rats, a single queen and several kings are responsible for all of the reproduction in the colony and all individuals are diploid (Figure 10.13). Although the workers in the group are capable of reproducing, they forgo reproduction in favor of caring for their younger siblings and taking care of the colony. Current research suggests that the subordinate males and females do not do this willingly. Instead, the dominant female harasses them, which causes them to be stressed. Stress reduces the levels of sex hormones in the subordinates and makes them less motivated to breed.

The Origins of Eusociality
From its distribution across a variety of distantly related species ranging from hymenopterans to mammals, it is clear that eusociality has evolved independently many times. Even within the hymenopterans, eusociality appears to have evolved several times. This indicates that eusociality was not due to a single mutation in an ancestor that caused all of the descendant species to be eusocial today. It is tempting to conclude that eusociality is caused by a haplodiploid sex-determination system, but many haplodiploid species are not eusocial, and some eusocial species—such as termites and naked mole rats—are not haplodiploid. Based on these observations, we are left to conclude that while being haplodiploid appears to favor the evolution of eusocial behavior by providing large indirect fitness effects when workers forgo breeding to help their sisters, it is not required for the evolution of eusocial behavior.
Throughout our discussion, we have focused on the importance of the coefficient of relatedness with the idea that the coefficient needs to be relatively high to favor eusociality. However, this equation could also be satisfied if the cost of forgoing personal reproduction is very low. If individuals that chose to leave a colony had a very low likelihood of surviving and setting up a new colony, then direct fitness would be very low. In naked mole rats, for example, some individuals leave the home colony to form new colonies, but most of these small new colonies do not persist beyond one year. As a result, the cost of forgoing reproduction—in terms of lost direct fitness—would be very small. When this cost is small, a large coefficient of relatedness is no longer required to favor eusocial behavior.
243
The origins of eusocial behavior are still actively debated. Many researchers have argued that it has been favored by the inclusive fitness of the altruistic sterile offspring. Others have argued that it has evolved to enhance the direct fitness of parents that force the offspring to forgo reproduction. Like many biological questions, continued research will likely provide new insights that will help us to understand eusocial societies.
Behavioral relationships among the social insects represent one extreme along a continuum of social organization, from animals that live alone except to breed to those that aggregate in large groups organized by complex behavior. Regardless of their complexity, all social behaviors balance costs and benefits to the individual, and the magnitude of these costs and benefits is often determined by the ecological conditions in which these behaviors exist. Like morphology and physiology, behavior is strongly influenced by genetic factors and therefore is subject to evolutionary modification by natural selection. The evolution of behavior becomes complicated when individuals interact within a social setting, and the interests of individuals within a population may either coincide or conflict. Understanding the evolutionary resolution of social conflict in animal societies continues to be one of the most challenging and important concerns of biology.
ECOLOGY TODAY CONNECTING THE CONCEPTS
HEN-PECKED CHICKENS

The social behavior of animals in the wild is inherently interesting, but understanding social behavior also has practical applications for domesticated animals. There are costs and benefits of raising domesticated animals in large groups. Raising animals such as pigs and chickens in high densities is more cost effective because it requires less space. However, as we have seen, living at high densities has downsides that include increased risks of disease transmission, increased competition, and increased fighting. For farmers who raise domesticated animals, raising animals in high densities translates into fewer healthy animals and smaller profits. However, during the past 2 decades, researchers with insights into group living have helped breed more productive domesticated animals. Some of the leading research has been conducted with chickens (Gallus gallus).
244
Chickens are well known for fighting when raised under crowded conditions. For example, if one chicken receives a small injury that causes a spot of blood on its feathers, other chickens will begin to peck at the spot, causing greater injury. In the process, some chickens will become dotted with blood spots and other chickens, in turn, will also peck them. This incessant behavior is the origin of the phrase “hen-pecked.” Fighting among chickens can reduce overall health and egg production, and can even cause death of the chickens. In fact, fighting can cause so much lost profit to poultry farms that there is a long-standing practice of cutting off the ends of chicken beaks to reduce injuries. Although beak trimming is effective at reducing fighting and cannibalism, it is a controversial practice and was banned in the European Union in 2011. This has motivated animal breeders to find alternative ways to reduce fighting among crowded domesticated animals.
The traditional method of breeding animals has been to select for those individuals with the best survival, growth, or other traits such as increased egg production. After decades of selection on traits, the amount of heritable variation in these traits is quite low, which makes it difficult to conduct further selection. In the mid 1990s, however, researchers discovered that they could cause large responses to selection if they based their selection on the best performing social groups of chickens living together rather than on just the best performing individual chickens. Groups of hens were caged together and cages with the highest survival were used to generate offspring for the next generation. At the start of the experiment, annual hen mortality was 68 percent. After six generations of selection for the best surviving groups, annual hen mortality dropped to 9 percent, which was similar to the mortality rate of hens that were raised alone with no opportunities to fight. Moreover, lifetime egg production increased dramatically, from 91 eggs per hen in the first generation to 237 eggs per hen by the sixth generation. The increase in egg production was caused by the hens living longer and the hens being in better physical condition, which allowed them to lay more eggs per day. These data suggest that selection for the best social groups could reduce fighting to a level that would eliminate the need to trim chicken beaks.
During the past 5 years, researchers have begun to understand why selecting on social groups of animals could cause such dramatic increases in animal production. Biologists have traditionally focused only on the heritability of traits for an isolated individual. However, the performance of an individual depends not only on the individual but also on the way it interacts with other individuals. For example, if an individual is particularly aggressive and lives in a group of other aggressive individuals, the group will spend more time fighting and more deaths will occur. In contrast, a social group with individuals who fight less with each other will have lower stress, better health, and a reduced mortality rate. Because social animals interact with conspecifics, we need to consider the heritability of traits not just when an individual lives alone but when an individual lives in a social group. In chickens, for example, the heritability of survival for an individual can be quite low, but the heritability of the trait when social interactions are included can be two to three times higher because social interactions affect survival. Similar results have been observed in other farmed animals including domesticated pigs (Sus scrofa) and mussels (Mytilus galloprovincialis). All of this suggests that by considering the social interactions of animals we can improve agricultural production of domesticated animals.
SOURCES: Muir, W. M. 1996. Group selection for adaptation to multiple-hen cages: Selection program and direct responses. Poultry Science 75: 447–458.
Wade, M. J., et al. 2010. Group selection and social evolution in domesticated animals. Evolutionary Applications 3: 453–465.
245