Populations fluctuate naturally over time
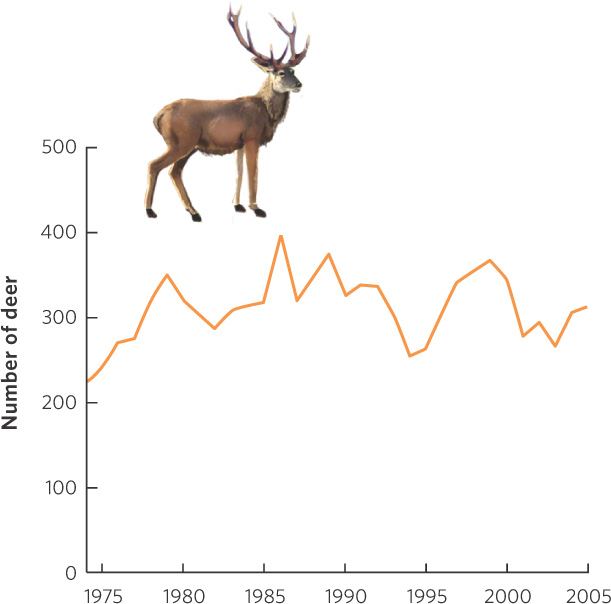
All populations experience fluctuations in size over time. There are many reasons for these fluctuations, such as changes in the availability of food and nesting sites, predation, competition, disease, parasites, and weather or climate. Despite these fluctuations, some populations tend to remain relatively stable over long periods. For example, researchers have studied fluctuations in a population of red deer—a close relative of the North American elk—on the Isle of Rum in Scotland for over 30 years. During that time, the population has been relatively stable, fluctuating between approximately 200 and 400 individuals, as shown in Figure 13.1.
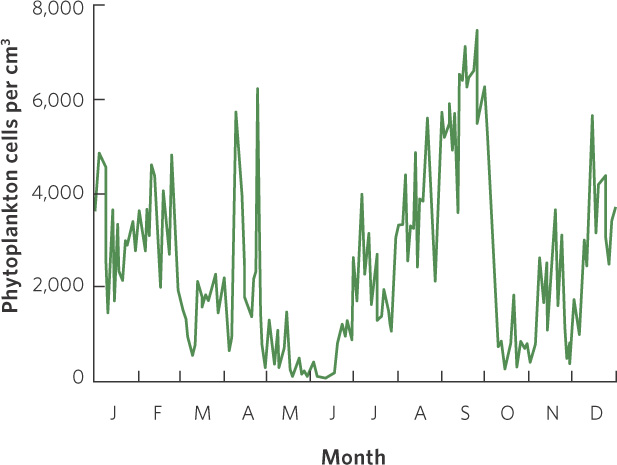
Other populations exhibit much wider fluctuations. In Lake Erie, for example, researchers monitored the number of waterborne algal cells, known as phytoplankton, throughout the year. As you can see in Figure 13.2, the algae fluctuated widely from nearly 0 cells per cm3 in June to more than 7,000 cells per cm3 in September. These wide fluctuations occurred across scales of days, weeks, and months.
297
Red deer populations are inherently more stable than algae. We can explain this by looking at differences in body size, population response time, and sensitivity to environmental change. Small organisms such as algae can reproduce in a matter of hours, which means that their populations respond very quickly to favorable and unfavorable environmental conditions. Their small bodies and the associated high surface-area-to-volume ratio cause them to be much more affected by unfavorable environmental changes, including abiotic conditions such as temperature. As a result, their rates of survival and reproduction can decline quickly. With larger animals that live for several years, a population at a given time includes individuals born over a long period, which tends to even out the effects of short-term fluctuations in birth rate. In addition, organisms with large body sizes can maintain homeostasis in the face of unfavorable environmental changes and therefore exhibit higher survival.
Age Structure Fluctuations
We have discussed population fluctuations over time by looking at surveys of population size. In some cases, however, we can detect fluctuations over time by examining a population’s age structure. When a certain age group contains an unusually high or low number of individuals, it suggests that the population experienced unusually high birth or death rates in the past.
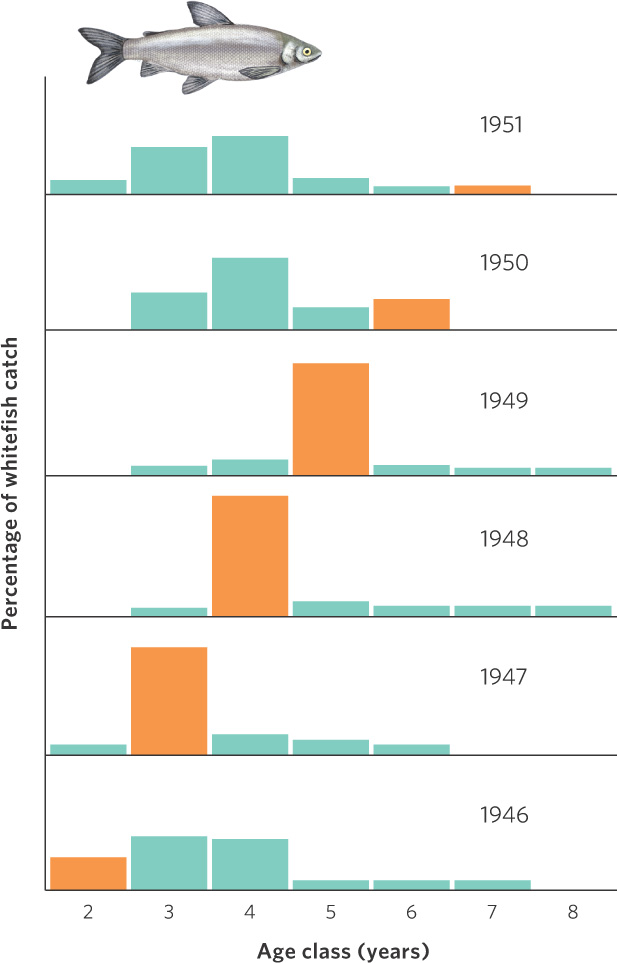
A classic example of age structure as an indication of population fluctuations comes from data of the commercial harvest of whitefish (Coregonus clupeaformis) in Lake Erie from 1945 to 1951. Biologists are able to determine the age of each fish by examining the scales of the harvested fish. As you can see in Figure 13.3, in 1947 there was a particularly large number of 3-year-old whitefish. This suggests that in 1944 the whitefish population experienced a very high amount of reproduction. This cohort continued to dominate the population’s age structure in subsequent years, with high numbers of 4-year-old fish in 1948 and 5-year-old fish in 1949. Even in 1950, the 6-year-old age class contained more individuals than had been observed in that age class in previous years.
The age structure of a forest is also readily analyzed. As you may know, the age of a tree can be determined by counting the number of rings in its trunk; under most circumstances, one ring is added each year. To examine how fluctuations affect the age structure of forest populations, researchers work at sites that have not been logged, because logging alters the age structure of the current tree population.
298
In one study, researchers investigated a site in Pennsylvania named Hearts Content, a small area of old-growth forest that had been protected from logging for more than 400 years. Researchers drilled into the trunks of trees to remove a sample of wood that contained tree rings. Using these samples allowed them to determine the age of each tree and, therefore, the time when each tree started its life.
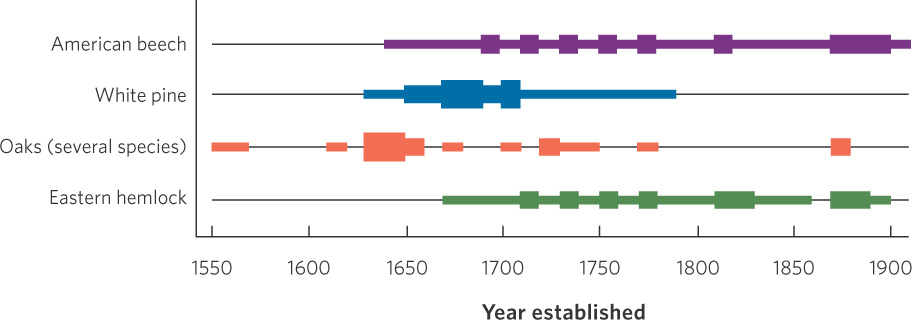
The data from this survey of trees is illustrated in Figure 13.4. The researchers found that in the 1500s the forest was largely composed of several species of oak trees. A fire and drought in the mid-1600s created openings in much of the forest. Following the fire, oak seedlings re-established themselves in the forest, although there was also an increase in the proportion of new white pines (Pinus strobis), which grow well under low-shade conditions. As the white pines grew large, however, they cast so much shade that new white pine seedlings had a hard time surviving, leading to a decline in the recruitment of white pine during the late 1700s. As a result, most white pine trees in the modern forest dated from the late 1600s and early 1700s. In contrast, American beech trees (Fagus grandifolia) and eastern hemlock trees (Tsuga canadensis) are very tolerant of high-shade environments; new individuals of these species began to grow as the white pines came to dominate and shade the forest. These two species continued to recruit new individuals over time, which caused beech and hemlock to have a population age structure that was more evenly distributed than that of white pine.
Overshoots and Die-Offs
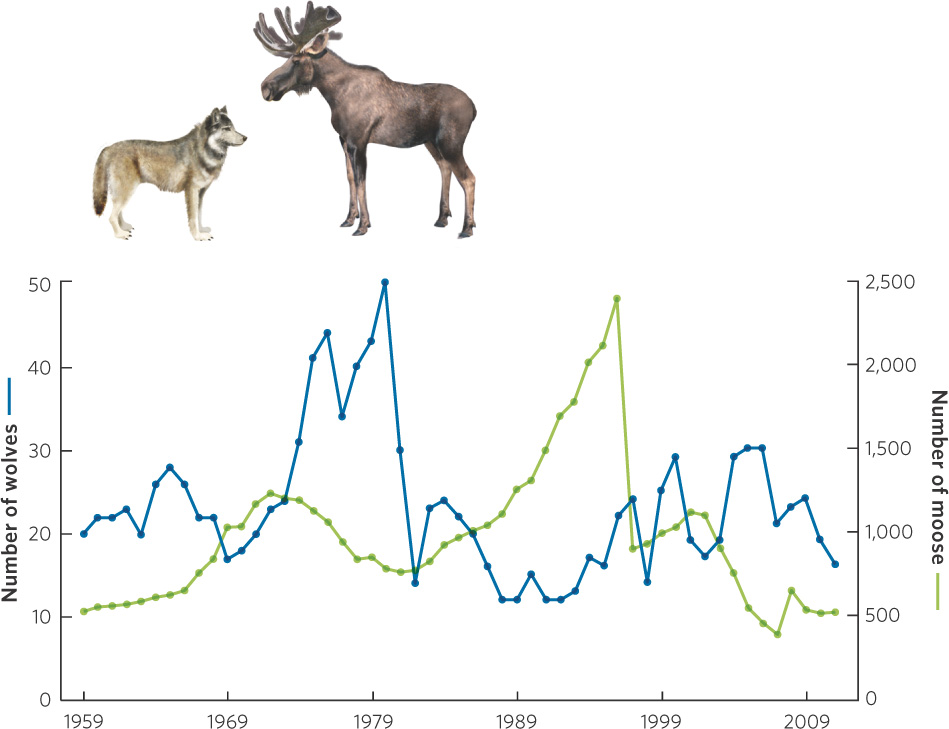
In Chapter 12, we saw that modeled populations experiencing density dependence initially grow quite rapidly but that this growth rate eventually slows as the population reaches its carrying capacity. However, populations in nature rarely follow a smooth approach to their carrying capacity. In many cases, they grow beyond their carrying capacity, a phenomenon ecologists refer to as an overshoot. An overshoot can occur when the carrying capacity of a habitat decreases from one year to the next. For example, if one year has abundant rainfall and the next year has a drought, the habitat will produce less plant mass in the second year. As a result, the carrying capacity for herbivores that rely on the plants for food will be reduced. A population that overshoots its carrying capacity is living at a density that cannot be supported by the habitat. Such populations experience a die-off, which is a substantial decline in density that typically goes well below the carrying capacity. Figure 13.5 illustrates an overshoot and die-off in a population of a hypothetical organism. The population begins small but grows at such a rapid rate that it exceeds the carrying capacity. Following the overshoot, the population experiences a rapid die-off to a point that is below the carrying capacity. We saw an example of this in the story at the beginning of this chapter that described wolves and moose on Isle Royale. Over time the moose exceeded their carrying capacity and the population experienced massive die-offs from starvation. You can see these data in Figure 13.6.
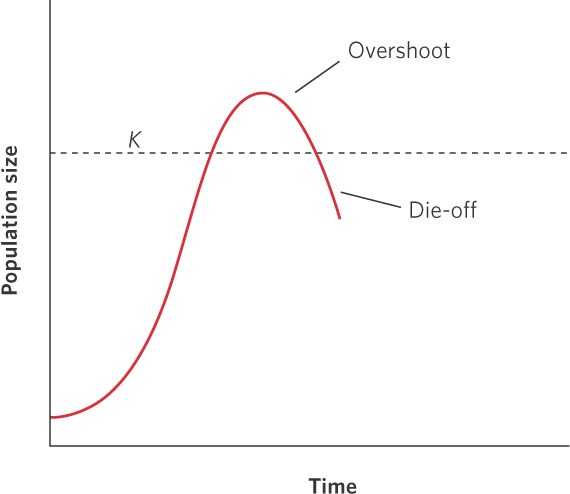
Overshoot When a population grows beyond its carrying capacity.
Die-off A substantial decline in density that typically goes well below the carrying capacity.
299
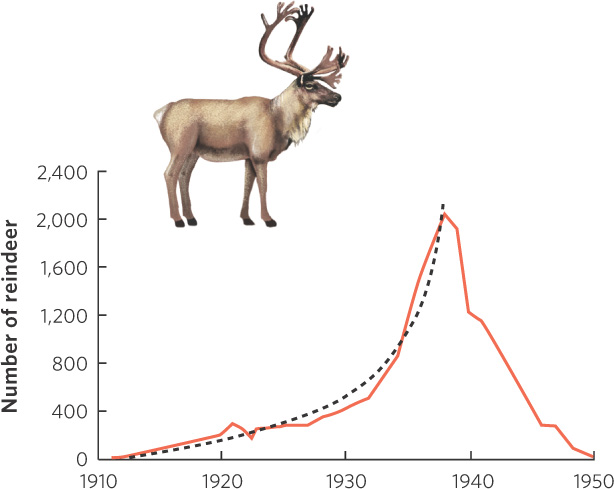
A natural experiment with reindeer in Alaska provides another good example of overshoots and die-offs. In 1911, the United States government introduced 25 reindeer to St. Paul Island, Alaska, to provide a source of meat for the local population. The island contained no predators of reindeer and the reindeer population quickly began to reproduce. The reindeer fed on a variety of items during the spring, summer, and winter, but they relied on lichens to get them through the winter. By 1938, the reindeer population had swelled to more than 2,000 individuals. As you can see in Figure 13.7, this rate of growth followed a J-shaped growth curve that we associate with exponential growth models. As the population grew in the early years, the lichens they consumed in the winter remained abundant. However, with continued growth of the reindeer population, the lichens became rare, which suggests that the reindeer had far exceeded the carrying capacity of the island. Following a peak in 1938, the reindeer population began to experience a massive die-off, probably from a combination of scarce winter food and unusually cold winters. In 1940 and 1941, the government culled several hundred reindeer in an attempt to reduce the size of the herd to get it closer to the island’s now much-reduced carrying capacity. Despite this effort, the population continued to decline and, by 1950, only 8 individuals remained. In 1951, 31 new reindeer were brought to the island to supplement the population. Since 1980, the population has been managed by the St. Paul Tribal Government, which maintains the population at just a few hundred animals. In 2012, the population was estimated to be 125 reindeer. The reindeer, maintained at a sustainable population size, can now continue to provide valuable and affordable meat to the local residents.
300
Cyclic Population Fluctuations
Population cycles Regular oscillation of a population over a longer period of time.
As we have just seen, populations can experience large fluctuations over time, including overshoots and die-offs. Over longer periods, some populations oscillate up and down in regular patterns known as population cycles.
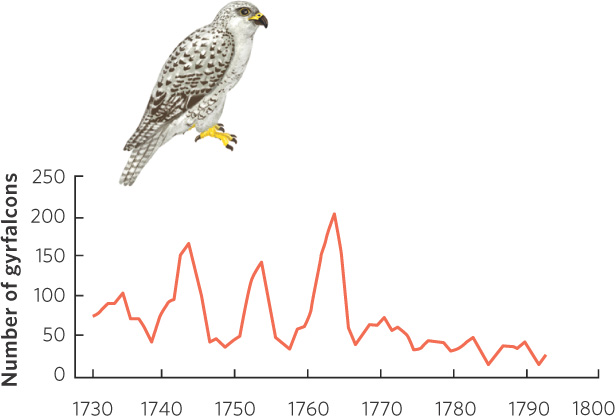
An interesting example of a population cycle comes from long-term records on the gyrfalcon. Falconry was a popular pastime among European nobility during the seventeenth and eighteenth centuries, and the gyrfalcon was especially prized because it is one of the largest and most beautiful falcons. During this time, Danish royalty imported gyrfalcons from Danish territories in Iceland where the falcons were trapped and then transported to Copenhagen. The Danish royalty would then present the falcons as diplomatic gifts to the royal courts of Europe. The governor of Iceland wrote the export permits, which provide a detailed historical record of gyrfalcon exports over several decades. As shown in Figure 13.8, the number of gyrfalcons exported from Iceland between 1731 and 1793 reveals 10-year cycles of abundance. After 1770, falconry became less popular and exports declined. Exports no longer exhibited regular fluctuations because the low demand for the birds could be satisfied even during years of low abundance.
In some cases, cyclic population fluctuations occur among species and across large geographic areas. In Finland, for example, biologists conducted annual surveys in 11 provinces to determine the abundance of three species of grouse: capercaillie (Tetrao urogallus), black grouse (Tetrao tetrix), and hazel grouse (Bonasa bonasia). After monitoring the birds for 20 years and graphing the data—as shown in Figure 13.9—they found that all three species experienced population cycles every 6 to 7 years. Moreover, these three species appear to exhibit high and low populations at the same time and this synchrony occurred across all of the provinces.
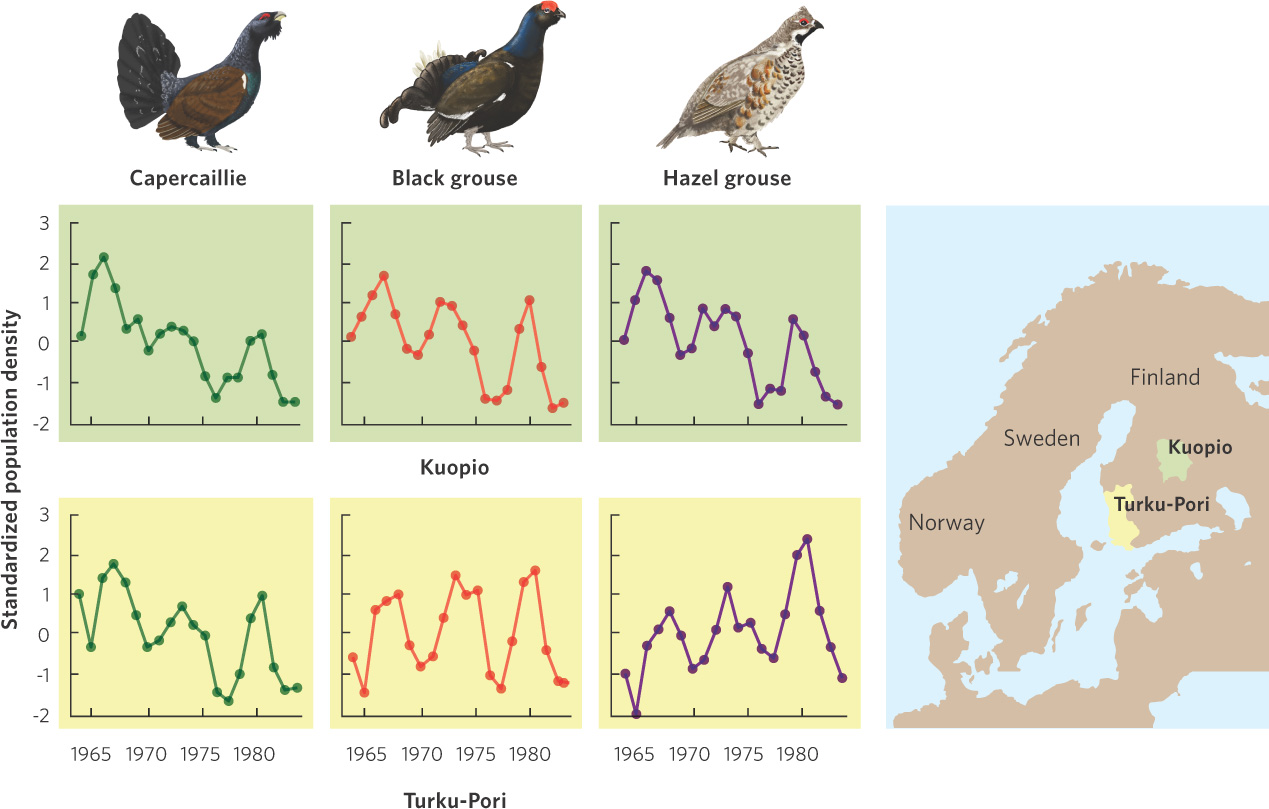
These examples demonstrate that some populations can exhibit remarkably regular fluctuations. The cause of such cycles, and of their synchrony, has been an interesting and persistent question in ecology. In subsequent chapters we will look at how species interactions also influence population cycles.