Communities respond to disturbances with resistance, with resilience, or by switching among alternative stable states
Communities commonly experience disturbances from both natural and human causes. Whatever the reason behind the disturbance, the key question is whether the community will be affected and, if it is, whether it will bounce back or become a substantially different community. To address this question, we need to consider the concepts of community stability and alternative stable states.
Community Stability
Community stability The ability of a community to maintain a particular structure.
As we have seen throughout this chapter, communities can experience large changes when the abundances of particular species change. Given enough time, most altered communities can rebound and resemble their original structure of species richness, composition, and relative abundance. The ability of a community to maintain a particular structure is known as community stability. There are two aspects of community stability, community resistance and community resilience. Community resistance is a measure of how much a community changes when acted upon by some disturbance, such as the addition or removal of a species. For example, we might measure community resistance after a predator is removed by looking at how much the abundance of herbivores increases. The smaller the change, the greater the resistance of the community to the disturbance. Community resilience is the time it takes after a disturbance for a community to return to its original state.
Community resistance The amount that a community changes when acted upon by some disturbance, such as the addition or removal of a species.
Community resilience The time it takes after a disturbance for a community to return to its original state.
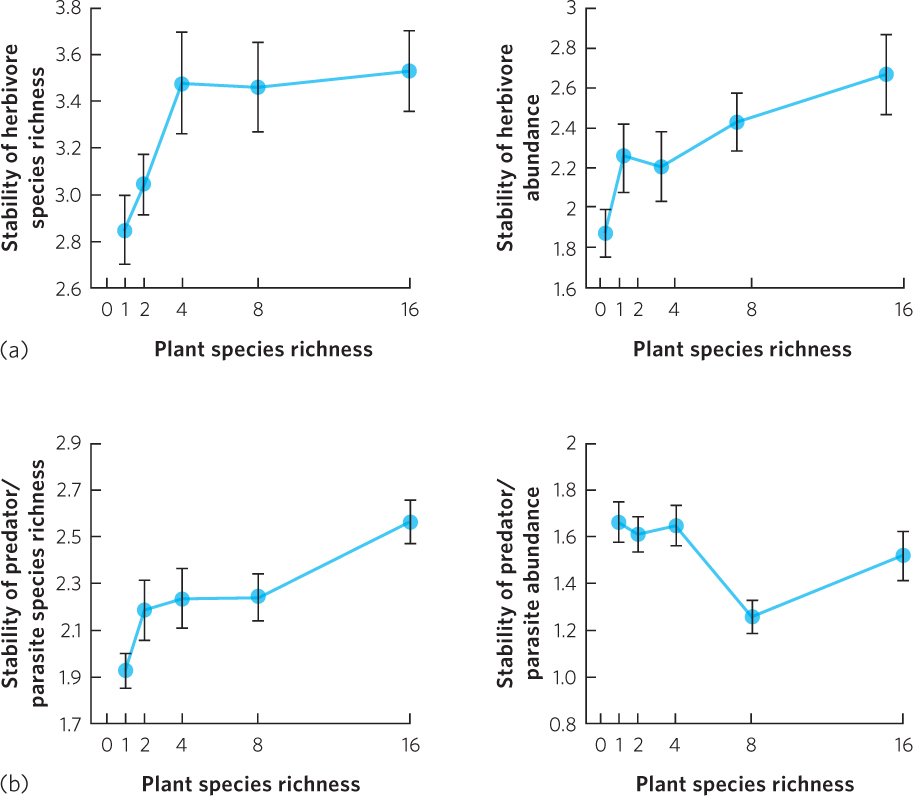
Does having more species help communities bounce back faster from disturbances? This is an interesting ecological question and a question of practical importance as human activities increasingly alter the species diversity of biological communities. One of the most extensive experiments on this question was conducted in grasslands where researchers manipulated the diversity of plant species in a Minnesota prairie ecosystem by 1, 2, 4, 8, or 16 species. For 11 years they monitored the abundances of more than 700 species of invertebrate herbivores and invertebrate predators and parasitoids. During the experiment, when the environment varied considerably, researchers asked which level of plant diversity provided the greatest community stability. In this study, community stability was measured as the amount of year-to-year variation in the abundance and species richness of the herbivores and predators.
In 2011, the researchers reported that increasing the number of plant species provided increased stability of the communities, as you can see in Figure 18.22. When they examined the herbivores, shown in Figure 18.22a, they found that an increase in plant species richness caused an increase in the stability of herbivore richness and herbivore abundance. When they examined the predators and parasitoids, shown in Figure 18.22b, they found that an increase in plant species richness caused an increase in the stability of richness, but there was no effect on the stability of abundance. Plant diversity led to increased stability of higher trophic levels because communities with high plant diversity provided a more consistent availability of food and habitat for the herbivores, predators, and parasitoids.
436
Alternative Stable States
Alternative stable state When a community is disturbed so much that the species composition and relative abundance of populations in the community change, and the new community structure is resistant to further change.
Sometimes when a community is perturbed, it does not bounce back to its original state. Rather, it settles into an alternative stable state, which occurs when a community is disturbed so much that the species composition and relative abundance of populations in the community change, and the new community structure is resistant to further change. For a community to move from one stable state to an alternative stable state typically requires a large disturbance, such as the removal of a keystone species or a dramatic change in the environment.
Moving between alternative stable states happens commonly where the prairie and forest biomes come together in the midwestern United States. During years of abundant rain, fire is suppressed and trees spread out into the prairie. Once trees have become established, they shade out many of the grasses and the grass species have little chance of returning. Moreover, the shade of the trees keeps the ground moist, which further reduces the likelihood of a future fire. During years of drought, however, fires are more common and large fires can kill trees, which favors the spread of grasses where trees once lived. Once grasses establish, it is difficult for trees to move into the prairie because grasses are well adapted to resprouting after fires, whereas trees are not.
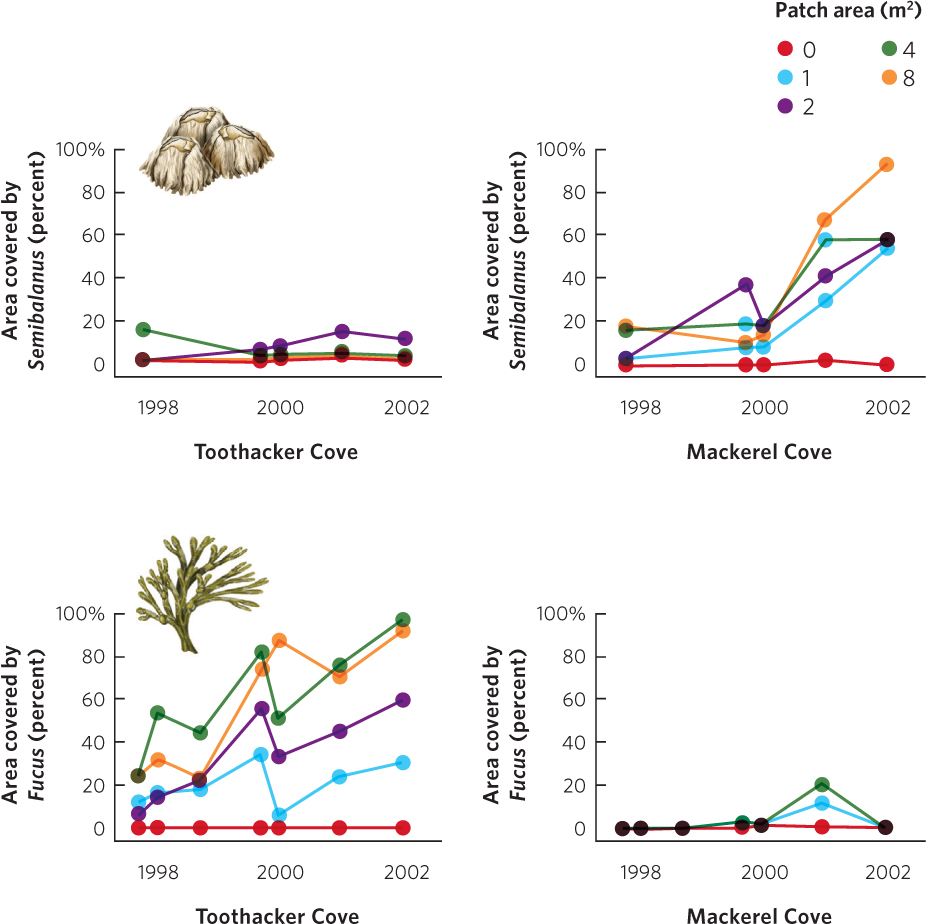
Alternative stable states also occur in aquatic environments. For example, intertidal communities on the coast of Maine are typically dominated by the brown rockweed algae Ascophyllum nodosum. During the winter, however, the scouring action of ice can scrape areas free of the rockweed algae and these areas become dominated either by another brown algae, Fucus vesiculosus, or by the barnacles Semibalanus balanoides. To determine whether these alternative communities are dependent on large disturbances, researchers simulated ice scouring on the coast by clearing areas of 1, 2, 4, and 8 m2. As shown in Figure 18.23, small scoured areas did not change to alternative communities, but the largest scoured areas did; the north-facing site, Mackerel Cove, became dominated by barnacles because it was more protected from large waves. The south-facing site, Toothacker Cove, became dominated by F. vesiculosus because the site was more exposed to waves. The switch to an alternative state occurred because A. nodosum is a poor disperser so it is slow to reach newly scoured sites compared to F. vesiculosus and barnacles. However, A. nodosum is a superior competitor for space, so over time A. nodosum may eventually be able to take over the open areas. The researchers found that the brown algae and barnacles were able to continue dominating sites for at least 20 years.
437
In this chapter we have seen that communities are composed of large numbers of interacting species that operate within food webs. The interconnected nature of food webs means that changes in the abundance of one species can potentially affect many other species in the community. In “Ecology Today: Connecting the Concepts,” we will see that understanding the connectedness of species was a key to discovering the effects of pollution on amphibians.
438
ECOLOGY TODAY CONNECTING THE CONCEPTS
LETHAL EFFECTS OF PESTICIDES AT NONLETHAL CONCENTRATIONS

Appreciating the connectedness of species in a food web can help us understand some of the effects our activities have on ecological communities. Pesticides, for example, provide important benefits in protecting crops and improving human health, and their widespread use has made them common in ecological communities. Maintaining pesticide concentrations below levels lethal to nonpest organisms is the key to ensuring that pesticides harm only pests and not other species. Pesticides are tested on a range of species in the laboratory to determine concentrations that cause death. However, laboratory tests do not consider the fact that species are part of a food web. Therefore, researchers have asked if a pesticide that is not directly lethal to a species can nonetheless have indirect impacts that cause mortality.
Some of the best-studied pesticides are those used to control terrestrial insects that damage crops as well as insects such as mosquitoes that carry infectious diseases. The most commonly applied insecticide is malathion, which impairs the nervous system of animals and is highly lethal to insects and other invertebrates. Based on single-species laboratory tests, scientists thought that concentrations found in nature were not lethal to vertebrate animals.
In a series of experiments, researchers examined the effects of malathion in large outdoor tanks that contained many of the components of natural wetlands including algae, zooplankton, and tadpoles (see Figure 1.18b). It had been long known that insecticides such as malathion were highly toxic to zooplankton, which are tiny crustaceans, but little was known about how large reductions in zooplankton might affect other species in the food web.
In one such experiment, researchers added tadpoles of wood frogs and leopard frogs (R. pipiens) to tanks and then manipulated malathion in one of four ways: no malathion to serve as a control, 50 parts per billion (ppb) at the beginning of the experiment, 250 ppb at the beginning of the experiment, or 10 ppb once per week. These manipulations allowed the researchers to ask whether single, large applications and multiple, small applications could alter the food web in ways that could affect the amphibians.
Soon after malathion was applied, the zooplankton populations declined to very low numbers in all of the tanks treated with the insecticide. This decline was not surprising since zooplankton were known to be quite sensitive to insecticides, but it set off a chain of events that ultimately affected the amphibians. Zooplankton feed on phytoplankton. When malathion was added and the zooplankton were killed, the phytoplankton populations rapidly grew over the next few weeks. This rapid growth, known as an algal bloom, caused the water to turn pea green.
439
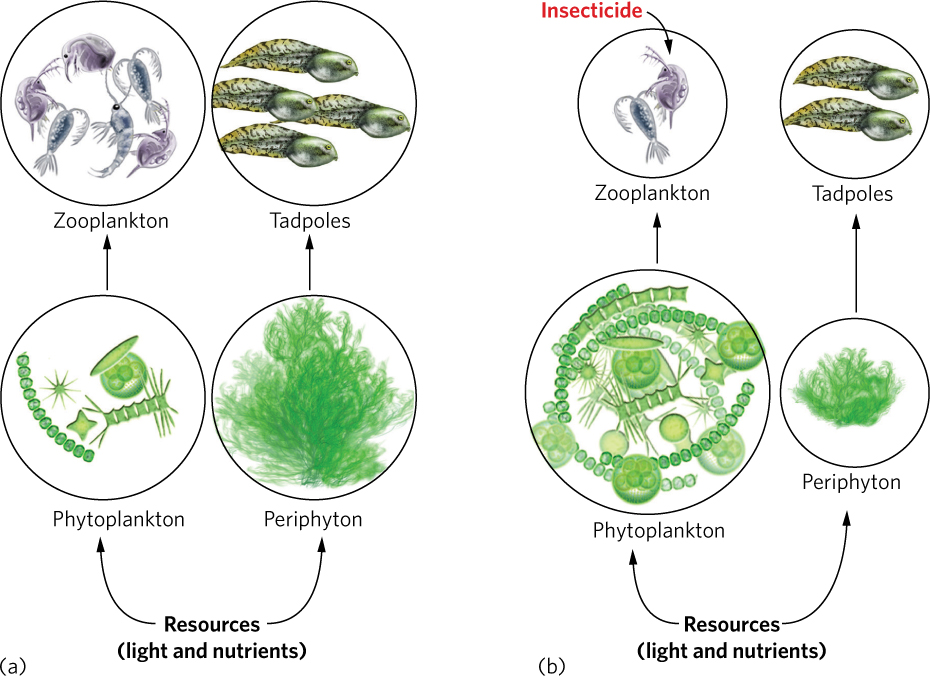
When the water turned green with phytoplankton, sunlight could no longer reach the bottom of the water column, where another group of algae, known as periphyton, grows attached to objects such as soil and leaves. The periphyton could not grow very well with less available sunlight, so within a few weeks after the start of phytoplankton bloom, the periphyton declined in abundance, especially in those communities that received 10 ppb of malathion every week. In this treatment, the decimated zooplankton populations could never exhibit resilience because the weekly additions of malathion continually reduced the rebounding populations.
Periphyton is the primary source of food for amphibians; without it, they cannot grow and metamorphose. Fortunately, the wood frogs develop rapidly. In the experiment, wood frogs metamorphosed after only 5 weeks, which meant that they were able to metamorphose before the bloom of phytoplankton caused a decline in their periphyton food source. As a result, wood frog growth and survival were not affected by the insecticide.
The leopard frogs, however, were not so fortunate. Because leopard frogs need 7 to 10 weeks of growth as tadpoles before they can metamorphose, they experienced the full effect of the declining periphyton when the insecticide was present. In fact, the leopard frogs had such stunted growth in the tanks with the weekly addition of 10 ppb of malathion that almost half of them failed to metamorphose before their aquatic environment dried in late summer.
This research demonstrated for the first time that a low concentration of an insecticide that is not directly lethal to amphibians can be indirectly lethal. The insecticide initiated a chain of events that moved through the food web and caused nearly half of the leopard frogs to die. Moreover, the smallest application of the insecticide, which was applied each week, had a much larger effect on the leopard frogs than single applications containing 25 times more insecticide.
These results underscore the importance of understanding ecology at the community level. Quantifying the insecticide’s direct effects on the amphibians suggested that the animals would not be harmed, but tests that incorporated a food web approach with indirect effects found that the insecticide could kill nearly half of the animals.
SOURCES: Relyea, R. A. 2005. The impact of insecticides and herbicides on the biodiversity and productivity of aquatic communities. Ecological Applications 15: 618–627.
Relyea, R. A., and N. Diecks. 2008. An unexpected chain of events: Lethal effects of pesticides on frogs at sublethal concentrations. Ecological Applications 18: 1728–1742.
440