Succession can occur through different mechanisms
Now that we have an idea of what succession looks like across a variety of communities, we can explore how succession actually occurs. In this section we consider the traits of early- and late-succession species, compare the different mechanisms of succession, and then examine succession studies to determine which mechanisms are common in ecological communities.
455
Traits Of Early- Versus Late-Succession Species
Early-succession and late-succession species possess different traits important to their performance. For example, pioneer species of terrestrial plants are typically better at dispersing seeds to newly created or disturbed sites. They produce many small seeds that are easily dispersed by wind or that stick to passing animals. These seeds can also persist in the soil for years and then germinate when a disturbance occurs. When they do germinate, these early-succession plants invest more into their shoots than their roots and grow rapidly and reproduce quickly. They are also typically quite tolerant of the harsh abiotic conditions that can exist in newly disturbed sites, including full sun and widely fluctuating temperatures and water availability. However, they are not tolerant of the high-shade conditions of late-succession plant communities.
In contrast, climax species produce a relatively small number of large seeds that disperse poorly; some simply drop to the ground whereas others are consumed by animals. The seeds have a relatively short viability and once they germinate they grow slowly, but their shade tolerance as seedlings and their large size as mature plants give them a competitive edge over early-succession species.
As succession progresses, we see a shift in the balance between adaptations that promote dispersal, rapid growth, and early reproduction, and adaptations that enhance competitive ability. Table 19.1 summarizes traits of early-succession and late-succession plants.
GENERAL CHARACTERISTICS OF EARLY-SUCCESSION AND LATE-SUCCESSION PLANTS
| TRAIT | EARLY SUCCESSION | LATE SUCCESSION |
|---|---|---|
| Number of seeds | Many | Few |
| Seed size | Small | Large |
| Mode of dispersal | Wind or stuck to animals | Gravity or eaten by animals |
| Seed viability | Long | Short |
| Root:shoot ratio | Low | High |
| Growth rate | Fast | Slow |
| Size at maturity | Small | Large |
| Shade tolerance | Low | High |
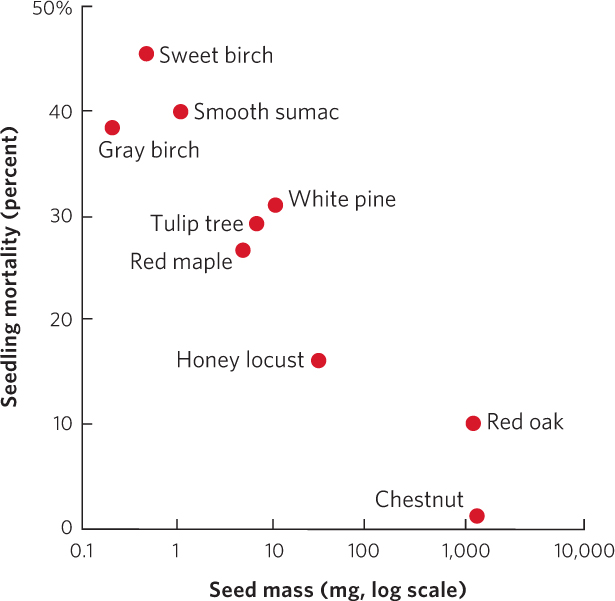
The traits of early-succession and late-succession plants are different because they face inherent trade-offs, similar to the life history trade-offs that we discussed in Chapter 8. To test for trade-offs between early-succession and late-succession species, researchers examined the mass of a single seed from each of nine species of trees. They raised the seeds under low-light conditions to simulate the high amount of shade in mature forests. After 12 weeks, they examined the mortality of the germinated seedlings, which you can view in Figure 19.11. The large-seeded species, which are common in old forests, experienced low rates of mortality under low-light conditions since large seeds provide their seedlings with ample nutrients to get started in the light-limited environment of the forest floor. In contrast, small-seeded species, which are common pioneer species that disperse well in large numbers, experienced high rates of mortality under low-light conditions. As a result, pioneer species cannot establish in mature forests.
456
Facilitation, Inhibition, and Tolerance
The ability to disperse and the ability to persist under existing abiotic and biotic conditions determine which species will appear in different seral stages during succession. Organisms that disperse well and grow rapidly have an initial advantage and therefore dominate the early stages of succession. Species that disperse slowly or grow slowly once they colonize an area typically become established later in succession. Early-succession species can also modify the environment in ways that affect whether late-succession species can become established. We must therefore consider whether a species has a positive, negative, or neutral effect on the probability of a second species becoming established. The mechanisms can be categorized as facilitation, inhibition, and tolerance.
Facilitation A mechanism of succession in which the presence of one species increases the probability that a second species can become established.
Facilitation is a mechanism of succession in which the presence of one species increases the probability that a second species can become established. Early-succession species do this by altering the environmental conditions of the site in a manner that makes it more suitable for other species to establish and less suitable for themselves. For example, alder shrubs, which are legumes, live in a mutualistic relationship with nitrogen-fixing bacteria in their roots, as we discussed in Chapter 17. This relationship produces additional nitrogen in the soil, which facilitates the establishment of nitrogen-limited plants such as spruce trees. Over time, the spruce trees eventually grow tall and cast a deep shade that is not a favorable environment for the alder shrubs.
Inhibition A mechanism of succession in which one species decreases the probability that a second species will become established.
Inhibition is a mechanism of succession in which one species decreases the probability that a second species will become established. Common causes of inhibition include competition, predation, and parasitism. That is, individuals of one species can inhibit those of other species by outcompeting them for resources, eating them, or attacking them with noxious chemicals or antagonistic behavior. Early in succession, inhibition can prevent movement toward a climax community, while late in succession, inhibition can prevent the pioneer species from colonizing and surviving. For example, in a mature forest in the northeastern United States, adult maple and beech trees cast a deep shade that prevents pioneer tree species from surviving.
Priority effect When the arrival of one species at a site affects the subsequent colonization of other species.
When inhibition occurs in a seral stage, the outcome of an interaction between two species depends on which species becomes established first. The arrival of one species at a site that affects the subsequent colonization of other species is known as a priority effect. We can see an example of the priority effect in the subtidal habitats of South Australia with bryozoans, a group of tiny invertebrate animals that live in colonies attached to rocks and that feed by filtering the water. If bryozoans become established first, they can prevent the establishment of tunicates and sponges—two other groups of filter-feeding animals that attach themselves to rocks. Sometimes the priority effect occurs because the first species to arrive has grown to a competitively superior adult stage whereas the second species to arrive is in the competitively inferior immature stage. For example, if a beech tree gets established in a forest and grows to an adult stage, it will cast deep shade on the ground below that prevents the seedlings of most other tree species from conducting sufficient photosynthesis to survive. In short, the mechanism of inhibition can make the path of succession dependent on which species arrives at the site first.
Tolerance A mechanism of succession in which the probability that a species can become established depends on its dispersal ability and its ability to persist under the physical conditions of the environment.
Tolerance is a mechanism of succession in which the probability that a species can become established depends on its dispersal ability and its ability to persist under the physical conditions of the environment. For example, species that can tolerate the stressful environmental conditions of early succession—such as low moisture or more extreme temperature fluctuations—can become established quickly and dominate early stages of succession. Similarly, plants that can tolerate high-shade environments can become established in forests that cast deep shade. These species do not alter the environment in ways that either facilitate or inhibit other species, but once a stress-tolerant species becomes established, it can be affected by interactions with other species. For example, superior competitors that arrive later will eventually replace the stress-tolerant species.
457
Tests for The Mechanisms Of Succession
For many years ecologists debated which of the three succession mechanisms were the most important determinants of the pattern of species replacement over time. Knowing the dominant mechanisms would allow scientists to predict how changes in communities proceed over time, especially since succession is often not a simple linear progression toward a climax community. To address this question, a great deal of research had to be done across a variety of biomes. Here we discuss two such studies: one conducted in an intertidal community and one conducted in a forest community.
Succession in Intertidal Communities
Sousa’s research in southern California showed that green algae prevented the colonization of red algae (see Figure 19.7.) This suggested that the succession of intertidal communities is determined by inhibition. However, the intertidal communities off the Oregon coast consist not only of large species of algae—known as macroalgae—but also of several common species of invertebrates that include the little brown barnacle (Chthamalus dalli), the common acorn barnacle (Balanus glandula), and several species of limpets, which are gastropods that consume algae. Observation of these communities indicated that they were dominated by the common acorn barnacle and a brown macroalgae, Pelvetiopsis limitata.
To determine how these communities came to be dominated by the two species, researchers scraped areas of rock clean and watched the succession of species over the subsequent 2.5 years. As illustrated in Figure 19.12a, the bare rocks were first colonized by the little brown barnacle. However, as more time passed, the larger acorn barnacle became established on the sites. As it became established, it slowly crushed the smaller little brown barnacle. Over time, the little brown barnacle became rare. Thus, the colonization of the acorn barnacle fit the tolerance model, but the decline of the brown barnacle fit the inhibition model. As the acorn barnacle became abundant, numerous species of macroalgae colonized the site and also became abundant. In fact, after 3 years, communities in the cleared sites closely resembled the control sites that had not been cleared.
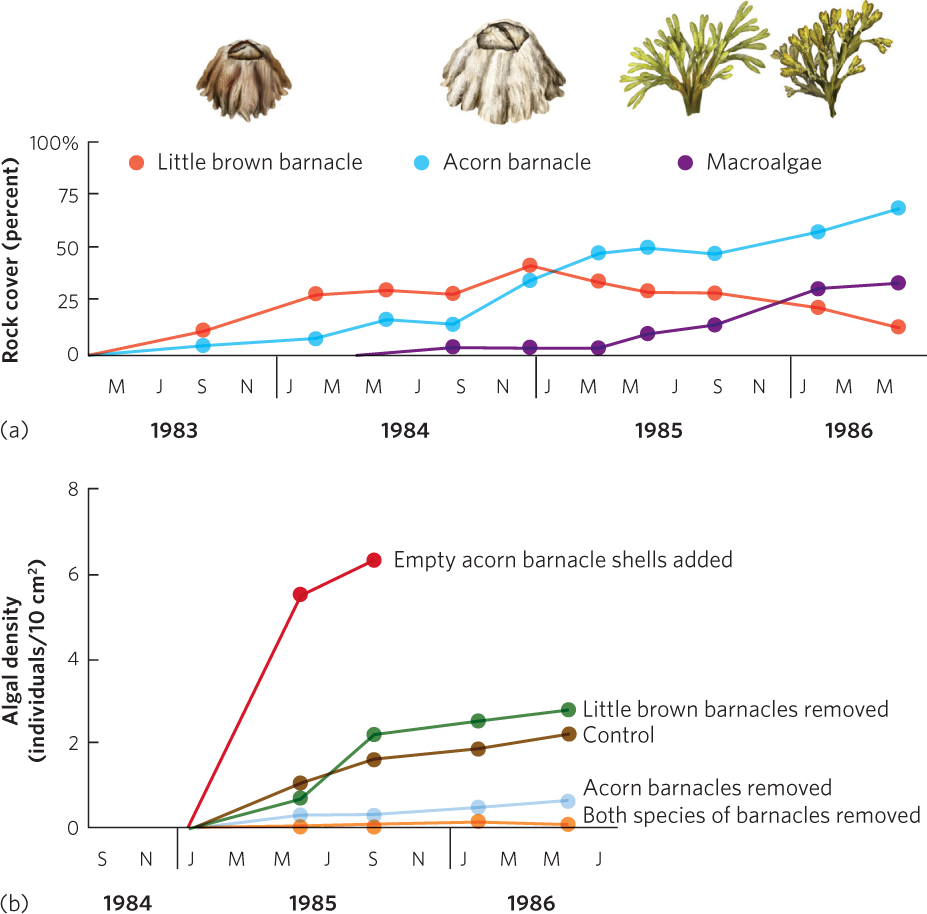
458
Which mechanisms of succession caused the increase of acorn barnacles and macroalgae? To answer this question, researchers conducted another experiment in which they removed different species from the community. They hypothesized that although the little brown barnacle would be the first to arrive, the acorn barnacle would eventually arrive and be a superior competitor for space. They also hypothesized that the presence of the acorn barnacles facilitated the colonization and survival of the macroalgae. The researchers began the experiment by once again scraping multiple sites clear of all organisms. Each site then received one of five manipulations: (1) no removal of any organisms, (2) removal of little brown barnacles, (3) removal of acorn barnacles, (4) removal of both species of barnacles, or (5) attachment of empty acorn barnacle shells. The final manipulation was used to test whether the mere physical presence of the acorn barnacle facilitated the colonization and growth of macroalgae by providing the macroalgae with protected crevices to which they could attach. The density of the macroalgae was then monitored for 2 years.
The results of this experiment are shown in Figure 19.12b. Starting near the bottom of the figure, you can see that the removal of the acorn barnacle, or both species of barnacles, caused a lower abundance of macroalgae than the control. In other words, the acorn barnacle facilitates the macroalgae. In contrast, the removal of the little brown barnacle had little effect on the density of the macroalgae. This confirmed that the little brown barnacle neither facilitates nor inhibits the macroalgae. However, the addition by researchers of empty acorn barnacle shells to an intact community that already included live acorn barnacles facilitated a large increase in macroalgae. Additional experiments revealed that the acorn barnacle facilitates the macroalgae by providing crevices where young macroalgae can attach to the rocks without being consumed by herbivorous limpets.
Collectively, these experiments in the intertidal zone indicate that the little brown barnacle persists because it is a good disperser that can rapidly colonize the frequently disturbed intertidal rocks and tolerate the conditions present on a bare rock. This is an example of tolerance. However, once the acorn barnacle arrives, it outcompetes the little brown barnacle, so inhibition also occurs later in succession. Finally, the succession of the macroalgae depends on the acorn barnacle, which is an example of facilitation.
Succession in Forest Communities
Like intertidal communities, forest communities can also exhibit a mixture of successional mechanisms. At Glacier Bay, for example, it was long hypothesized that each seral stage facilitates species in the subsequent stage. This made sense because later seral stages had soils containing greater organic matter, more nitrogen, and more moisture. If facilitation were the most important mechanism of succession, we would expect that a given species would grow and survive well in a seral stage where it dominates but have a difficult time growing and surviving in earlier stages.
To test this hypothesis, a group of researchers planted spruce seeds and seedlings in four seral stages of Glacier Bay: an early-succession pioneer stage containing lichens, mosses, and herbs; a low-shrub stage dominated by mountain avens (Dryas drummondii); an alder stage dominated by dense thickets of tall alder shrubs (Alnus sinuate); and a spruce stage dominated by spruce trees. You can see the results in Figure 19.13a. When spruce seeds were planted in bare plots within each seral stage, germination of these seeds was high in the pioneer stage, low in the low-shrub and alder stages, and high in the spruce stage. When spruce seedlings were planted in plots within each seral stage, seedling growth was high in the pioneer and low-shrub stages, none survived in the alder stage, and growth was low in the spruce stage, as shown in Figure 19.13b. In short, the spruce seeds and seedlings were capable of germinating and growing quite well in the earliest seral stages at Glacier Bay, yet spruce was a rare plant in these early stages. Moreover, whereas spruce seeds and seedlings grew well in the low-shrub and alder stages, seedling growth was inhibited in the spruce stage.
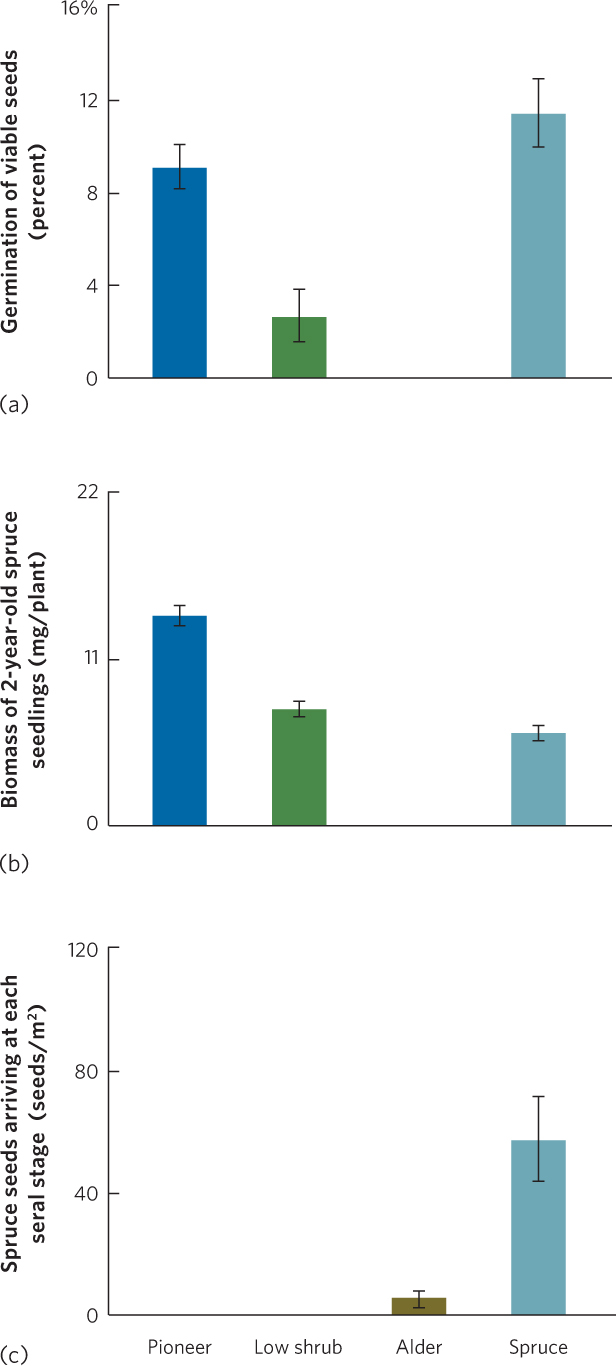
These observations refuted the hypothesis that each seral stage facilitates the next stage, but it led researchers to question why spruce trees were not common in the earlier stages. They decided to measure how many spruce seeds were dispersed by wind to each site. They discovered that very few spruce seeds arrived to colonize the pioneer and low-shrub stages. There were many more spruce seeds dispersed into the alder stage and even more dispersed into the spruce stage, as shown in Figure 19.13c. The researchers concluded that the dominance of spruce in later seral stages had little to do with facilitation or inhibition, but was due to differences in how many dispersing spruce seeds arrived in each location.
459