Succession does not always produce a single climax community
Ecologists have traditionally viewed succession as proceeding through a series of stages that end with a climax community, which remains constant over space and time unless another major disturbance occurs. As we saw in the case of sand dune succession around Lake Michigan, a site can follow different pathways of succession and end up at the same climax community. However, the species composition of a climax community can still exhibit variation over space and time within a given biome. The climax community can also be an ephemeral condition because disturbances can wipe it out. In this section, we will examine how climax communities can change over time and how their composition varies along environmental gradients. We will also investigate how alternative climax communities can be favored in the presence of different soils and by disturbances of different sizes and intensities.
Changes in Climax Communities Over Time
When succession occurs in a community, we typically observe changing environmental conditions and a progression from small life forms to large life forms. For example, primary succession on land begins with lichens and mosses and progresses to grasses and herbs. When sufficient moisture is available, as is the case in eastern North America, succession can continue to a stage that includes large trees. As succession occurs, the abiotic conditions are rapidly altered; areas with trees have less light at ground level, lower ground temperatures during hot summer days, and higher soil moisture. However, once a point is reached where the community contains the largest plants it can support, changes in environmental conditions occur more slowly. As a result, the changes in the community become less dramatic once the climax community develops.
When environmental conditions become relatively stable, the composition of plant species that dominate the community also becomes relatively stable. However, the species found in a climax community can continue to change. For example, northern deciduous forests have a climax community dominated by large trees, but the composition of large tree species can slowly change over time. Initially these large trees are mostly oak, hickory, and tulip poplar (Liriodendron tulipifera). However, over time the dominant species can change to sugar maple and beech. In an old-growth forest in Pennsylvania, researchers surveyed the adult trees that lived in the canopy and the sapling trees that lived in the understory. After surveying the trees, they calculated the importance value of each tree, which incorporates both the abundance and total area of the trunks for each species. As you can see in Figure 19.14, the researchers discovered that the canopy contained a variety of species with high importance values that included sugar maple, American beech, tulip poplar, several species of oaks, and hickories.
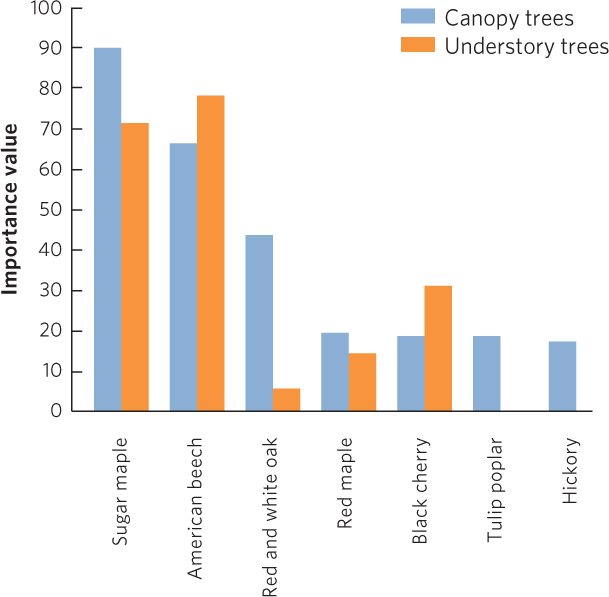
460
In contrast to this adult distribution, there were few oaks and no tulip poplars or hickories in the understory. The oaks, tulip poplars, or hickories were not in the understory because these species are not very tolerant of the deep shade that occurs in a mature forest and they are also susceptible to browsing by deer. In comparison, maple and beech trees are very tolerant of shade and less susceptible to deer browsing. As a result, maple and beech trees can survive and grow in the understory of large trees while other species cannot. Over time, the climax community of large trees will experience a gradual shift in the composition of the dominant species; as the current canopy species gradually die, there will be no younger oaks, tulip poplars, or hickories to replace them. In short, the composition of a climax community of large trees in a northern deciduous forest can continue to change over time.
Changes in Climax Communities Over Space
We have seen that the composition of a climax community can vary over time. Climax communities can also vary in composition as one moves along environmental gradients. For instance, in the 1930s plant ecologists described the climax vegetation of much of Wisconsin as a sugar maple–basswood (Tilia americana) forest. However, they later determined that the climax forest exhibited differences in various locations around the state. In the southern part of the state, beech trees were more common while in the north, birch, spruce, and hemlock trees were more common. In the drier regions bordering prairies to the west, oaks became prominent. On drier upland sites, quaking aspen, black oak, and shagbark hickory—long recognized as successional species on moist, well-drained soils—came to be accepted as climax species.
Transient Climaxes
Transient climax community A climax community that is not persistent.
Sometimes a climax community is not persistent, a phenomenon known as a transient climax community. A transient climax occurs when a site is frequently disturbed so that the climax community cannot continue to perpetuate itself. A common example of a transient climax occurs in small wetlands, sometimes known as vernal pools, which fill with water each spring and then either dry up in summer or freeze solid in winter (Figure 19.15). Although drying and freezing events eliminate most species that comprise a pond community, some species have resting stages and persist in the soil until the wetland refills again in the spring. For example, many species of zooplankton produce resting eggs that can persist in the dry bottom of a wetland and then hatch when the wetland fills with water again. Similarly, some species of snails can aestivate, as discussed in Chapter 4, which allows them to live under the soil surface of the dry pond with their metabolic processes largely shut down. When the water returns, plants, animals, and microbes come back to life from their resting stages. Many other species that live on land as adults, such as frogs, salamanders, aquatic beetles, and dragonflies, lay their eggs in the wetland. In this way, the community once again starts the process of succession, only to be destroyed by summer drying or winter freezing.

461
Creating Gaps in a Climax Community
Sometimes climax communities contain species that are not considered climax species. These species fill relatively large gaps created by small-scale disturbances in an area. In mature forests, for example, adult trees eventually die and leave a gap in the canopy that lets in sunlight (Figure 19.16). If the gap is not large, the surrounding branches of neighboring trees will likely grow in and close the gap. However, if the gap is large, the area of intense sunlight provides local conditions favoring species from earlier seral stages that have widely dispersed seeds and an ability to grow rapidly under high sunlight conditions. As a result, a mature forest that contains mostly climax species of trees can also contain a few early-succession trees. Gaps can occur in a variety of terrestrial and aquatic biomes. For example, in Chapter 18 we saw how gaps of increasing size in the intertidal zone favored the formation of alternative stable states (see Figure 18.23).

462
Climax Communities Under Extreme Environmental Conditions
As we have seen throughout this chapter, the composition of a climax community is determined by the environmental conditions that develop over time, including temperature, light, nutrients, and moisture. In some areas, however, additional environmental conditions also play a role, such as in communities affected by fire or grazing.
Fire-maintained climax community A successional stage that persists as the final seral stage due to periodic fires.
Biomes in which fires occur at regular intervals favor the persistence of fire-tolerant species. For example, in Chapter 16 we saw that the pine forests in the southeastern United States experience periodic fires that kill oak trees and other species of broadleaf trees, but not pine trees. In fact, some species of pines do not even release seeds from their cones unless triggered by the heat of a fire. After a fire, pine seedlings grow rapidly because there is little or no competition from other understory species. As a result, forest succession reaches a climax that is dominated by pine trees. When a successional stage persists as the final seral stage due to periodic fires, we call it a fire-maintained climax community.
The chaparral vegetation found in California is another community that has a fire-maintained climax. As we discussed in Chapter 6, the California chaparral is an example of the woodland/shrubland biome, which has cool, wet conditions in the winter and hot, dry conditions in the summer. As a result, plants can produce thick layers of detritus that become very susceptible to fires during the dry summers.
Grazer-maintained climax community When a successional stage persists as the final seral stage due to intense grazing.
When a successional stage persists as the final seral stage due to intense grazing, we call it a grazer-maintained climax community. Grazers can create a different climax community because they preferentially consume the most palatable plants and do not consume the less palatable or better-defended plants. In the dry grasslands of Arizona, for example, intense cattle grazing can kill or severely damage many species of grass and leave behind less palatable plants such as mesquite and cactus (Figure 19.17).

In western North America, cattle grazing allows invasion by cheatgrass (Bromus tectorum), a grass native to Europe, Asia, and North Africa that was introduced to North America in the late 1800s. Cheatgrass is able to colonize when grazing removes much of the competing grasses. Once it establishes, the detritus it produces is quite susceptible to fire. The grazing promotes the invasion of cheatgrass and the repeated fires that occur promote its long-term persistence. In this case, the change in climax community is maintained by a combination of grazing and fire.
Throughout this chapter, we have discussed how communities change over time after a major disturbance. We have emphasized that communities are always changing, both when we move from early to late seral stages and even when a community appears to have attained a climax state. By observing both what happens after a disturbance and the underlying mechanisms that cause the changes, we obtain new insights into the processes that regulate the structure of communities. It is also important to remember that the climate that influences succession is also changing, which means that succession toward a climax stage is actually heading toward a perpetually moving target.
463
ECOLOGY TODAY CONNECTING THE CONCEPTS
PROMOTING SUCCESSION ON A STRIP MINE

Coal has long been a major source of energy in North America and Europe. For several centuries humans have mined coal near the surface of the ground by removing the upper layers of soil and rock and then digging out the coal underneath. This process, known as strip mining, surface mining, or mountaintop-removal mining, is an efficient way of mining coal, but these mines eventually run out of coal and leave an immense barren landscape. Many areas of the world require the coal mining industry to return the barren land to a more natural condition.
Since the ground may have little or no organic soil remaining after strip mining, it is a challenge to manipulate the succession of communities to reclaim the stripped land. Prevention of soil erosion is critical and can be accomplished by rapidly colonizing the area with plants that will hold the soil and ultimately undergo succession in a way that promotes species richness and improves the functioning of the ecosystem. To promote succession on former mined areas, we need to understand how it works by observing the succession changes in plant communities.
In Spain, researchers examined a chronosequence of 26 strip mines in the northern region of the country that had been abandoned for 1 to 32 years. The mines all had similar soils and climate. When the researchers examined changes in species richness, they found that mines abandoned for 1 year had 8 plant species and that species richness peaked at 28 species after 10 years of being abandoned. This peak was caused by the persistence of some of the pioneer species combined with the colonization by later succession species. After 10 years, species richness exhibited a gradual decline; mines abandoned for 32 years had only 7 to 8 species. As with other communities discussed in this chapter, most of the species in the young abandoned sites were annual wind-dispersed plants. These species were able to tolerate the harsh conditions of the newly abandoned mine. After 10 years there was an increase in the number of perennial herbs, and woody plants started to colonize the area. These included nitrogen-fixing plants that contributed large amounts of organic matter and nitrogen to the soil, which facilitated subsequent woody plants, including trees that began to dominate after 20 years. These changes correlated with increased nitrogen in the soil, which suggested that the plants that formed the earliest communities facilitated the colonization and growth of later species.
Understanding how succession proceeds helps scientists develop recommendations for speeding up succession. For example, researchers in Germany examined how succession would be affected under three different manipulations: let the site naturally undergo succession; sow the area with a mixture of herb and grass seeds; or cover the area with newly cut hay, which helps to reduce erosion and provides the seeds of dozens of herbs and grasses contained in the soil. After 4 years, the plots experiencing natural succession had only 35 percent plant cover while the plots treated with either sown seeds or hay had more than 80 percent plant cover. These substantial differences in plant cover affected soil erosion; plots with sown seeds or green hay had channels that had eroded less than 5 cm deep while plots undergoing natural succession had channels that had eroded up to 1.5 m deep. The plots experiencing natural succession also had lower species richness during the first year and a lower index of similarity, although the richness of the three plots converged by the end of the 9-year experiment. Collectively, these data confirm that while primary succession will naturally occur on abandoned strip mines, we can use our knowledge of succession to speed up the process and rapidly move a barren landscape toward a much more natural community. However, as we have learned throughout this chapter, terrestrial communities may require centuries of succession after a disturbance such as strip mining before they can approximate the communities that were originally present.
464
SOURCES: Alday, J. G., et al. 2011. Functional groups and dispersal strategies as guides for predicting vegetation dynamics on reclaimed mines. Plant Ecology 212: 1759–1775.
Baasch, A., et al. 2012. Nine years of vegetation development in a postmining site: Effects of spontaneous and assisted site recovery. Journal of Applied Ecology 49: 251–260.