On a global scale, biodiversity is highest near the equator and declines toward the poles
We have examined the local and regional processes that can affect the number of species in a given location on Earth. However, patterns of biodiversity also exist at the global scale. One of the most striking patterns is that species richness of all taxa combined is highest near the tropics and declines toward the poles. For example, a hectare of forest typically has fewer than five species of trees in boreal regions, 10 to 30 species of trees in temperate regions, and up to 300 species of trees in tropical regions. These latitudinal trends in diversity are pervasive and extend even to the oceans. In this section, we will explore the patterns of diversity within and across latitudes. We will then discuss the two general hypotheses for these patterns of biodiversity and explore three important processes.
Patterns of Diversity
In the Northern Hemisphere, the number of species in most groups of animals and plants increases from north to south. For example, when a researcher counted the number of mammal species that one could find in blocks of habitat that were 241 km on a side, he found that there were fewer than 20 mammal species per block in northern Canada but more than 50 species per block in the southern United States. You can view his data in Figure 22.21a.
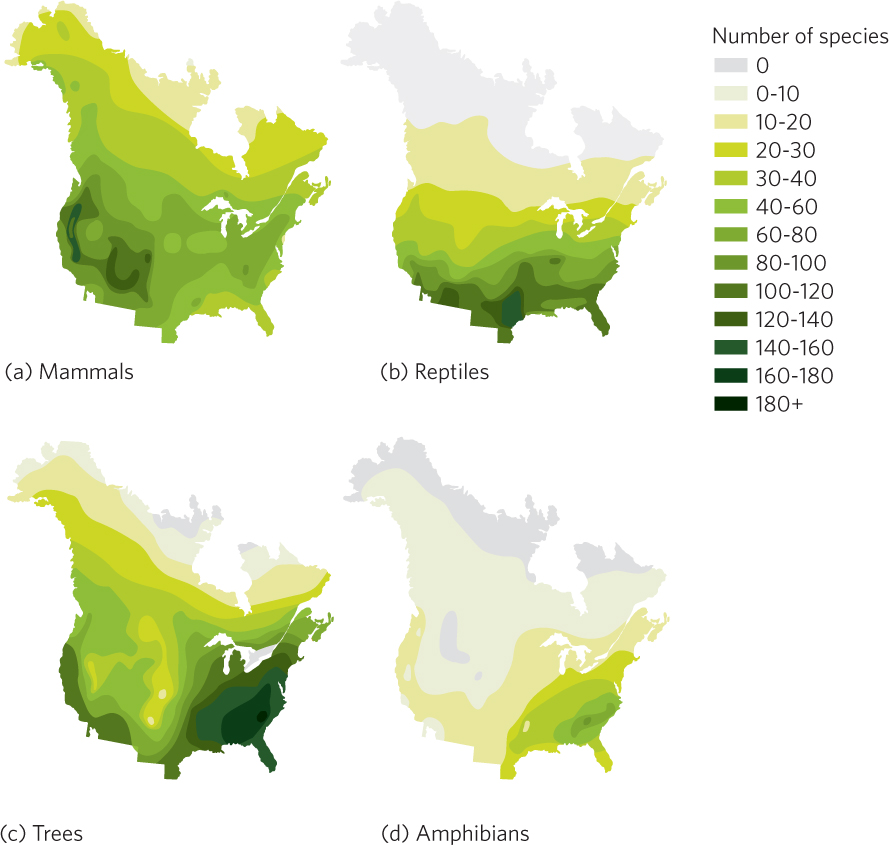
The number of mammals also increases as we move from east to west in North America. For instance, there are typically 50 to 75 species per sample block in the east whereas there are 90 to 120 species per block in the west. Such a pattern is likely due to the greater amount of habitat heterogeneity in the extensive mountain ranges of western North America. This greater heterogeneity of environments in the west apparently provides suitable conditions for a greater number of species, similar to the pattern we saw for plants in the Cape Floristic Region at the beginning of this chapter.
The pattern of species richness for breeding land birds in North America resembles the pattern for mammals, but the patterns for reptiles, trees, and amphibians are strikingly different, as you can see in Figures 22.21b–d. The richness of reptile species declines fairly uniformly as temperature decreases toward the north. However, trees and amphibians are more diverse in the moister eastern half of North America than in the drier, more mountainous western regions.
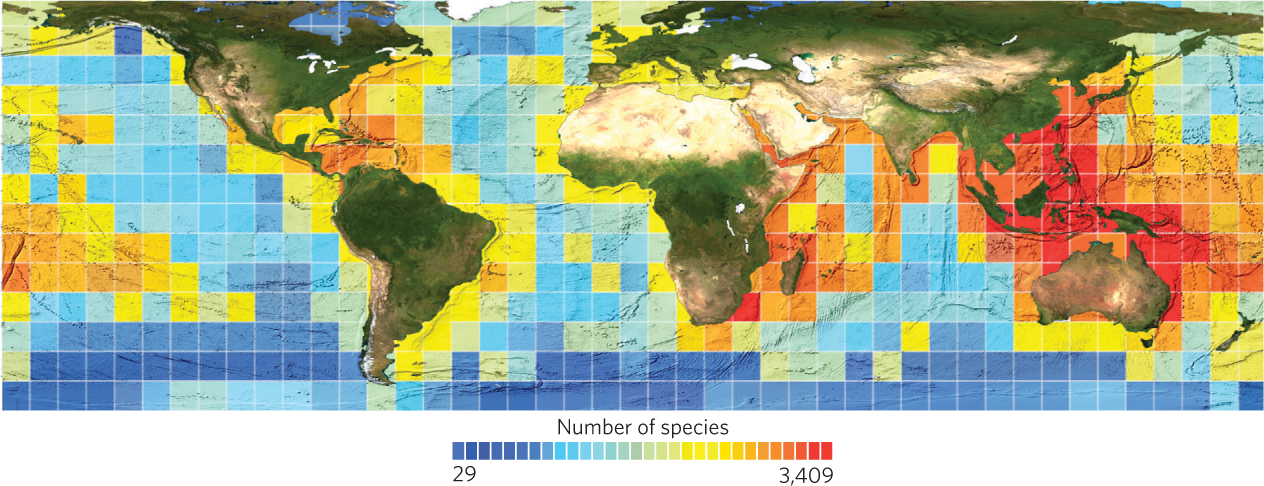
The general pattern of increased species richness at lower latitudes that we observed for trees and vertebrate animals also holds true for the oceans. For example, in 2010 researchers compiled millions of records of marine organisms around the world including whales, dolphins, fish, and corals. When they examined the global patterns of diversity using sample blocks of 880 km on a side, as shown in Figure 22.22, they found that the highest diversity occurred in the tropics including Central America and Southeast Asia whereas the lowest diversity occurred near the poles. Within a given latitude, they also found a higher diversity near the coasts and lower diversity in the open ocean.
531
Processes that Underlie Patterns of Diversity
For more than a century ecologists have investigated reasons for the decline in species richness as we move from the equator to the poles. Historically, ecologists have considered two general hypotheses. According to one hypothesis, species are continually created over time and without limit. Because the world’s current temperate and polar regions experienced repeated advances and retreats of glaciers during the Ice Age, species in these regions have been eliminated or driven toward refuges closer to the equator. In contrast, because the world’s tropical regions did not experience glaciation, habitats in these areas have remained stable much longer and so have had more time to accumulate species.
532
The second hypothesis proposes that the number of species reflects an equilibrium between the processes that create new species and the processes that drive species extinct, similar to the state of equilibrium described by the theory of island biogeography. According to this hypothesis, the higher number of species in the tropics is the result of higher rates of speciation or lower rates of extinction there compared to the temperate and polar regions. Similarly, variation in the number of species as one moves across a given latitude should also reflect an equilibrium between processes that create new species and destroy existing species. We will now consider three processes that play substantial roles in determining the number of species that occur in an area: ecological heterogeneity, solar energy and precipitation on land, and water temperatures in the oceans.
The Role of Ecological Heterogeneity
At any given latitude, we find more species in areas where there is greater ecological heterogeneity, such as heterogeneity in the soils and plant life. For example, grasslands contain vegetation that is less heterogeneous in growth form than shrublands or deciduous forests. Surveys of breeding birds in North America find an average of six species in grasslands, 14 species in shrublands, and 24 species in floodplain deciduous forests. You may recall that we discussed a similar phenomenon in Chapter 18 when we looked at the positive relationship between the diversity of vegetation height and the diversity of birds (see Figure 18.13). This same pattern has also been observed in lizards of the southwestern United States, where lizard diversity is associated with vegetation diversity, and in the plants of the Cape Floristic Region of Africa, where plant diversity is associated with the heterogeneity of the soils.
Although more productive terrestrial habitats tend to have more species, habitat heterogeneity can also play a role in determining species richness when two habitats have similar levels of productivity. For instance, habitats with less variation in vegetation growth form, such as grasslands, have fewer animal species than habitats with similar productivity but more variation in vegetation. This principle also applies to plants. Marshes are highly productive but have a relatively uniform landscape and thus contain relatively few species of plants. While desert vegetation is less productive than marsh vegetation, the greater heterogeneity of the desert landscape provides room for more species of plants (Figure 22.23).

The Role of Solar Energy and Precipitation on Land
Potential evapotranspiration (PET) The amount of water that could be evaporated from the soil and transpired by plants, given the average temperature and humidity.
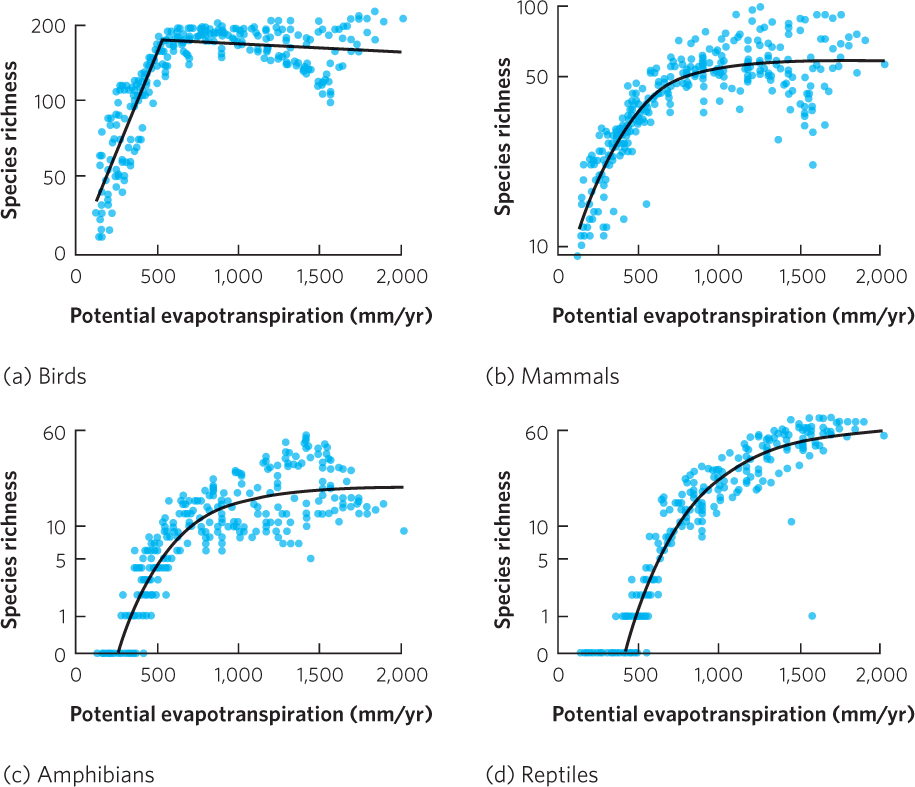
The number of species found at any location as one moves from the poles to the tropics is positively correlated to the amount of solar energy and precipitation at that location. Ecologists have combined solar energy and precipitation into a measure of potential evapotranspiration. Potential evapotranspiration (PET) is the amount of water that could be evaporated from the soil and transpired by plants, given the average temperature and humidity. Because PET integrates temperature and solar radiation, it provides an index to the overall energy input into the environment. As you can see in Figure 22.24, PET correlates with the species richness in North American vertebrates relatively well. In each group of vertebrates, the initial rise in PET is associated with a rise in species richness, which reflects increasing diversity from north to south within the continent. However, species richness eventually plateaus at very high levels of PET. These high PET levels occur as we move from east to west at middle latitudes where the increasing temperature fails to improve the capacity of the environment to support additional vertebrate species. In the arid western parts of the continent, for instance, increasing temperature eventually becomes a stressor.
533
Energy–diversity hypothesis A hypothesis that sites with higher amounts of energy are able to support more species.
The correlations between PET and species diversity for terrestrial vertebrates has given rise to the idea that there is a causal relationship between the two variables. This hypothesized causal relationship is known as the energy-diversity hypothesis, which states that sites with higher amounts of energy are able to support more species. Higher amounts of energy would also support higher abundances of individuals from each species, which should reduce the rate of extinction. Moreover, higher energy input might accelerate the rate of evolutionary change and thereby increase the rate of speciation. While these ideas are attractive, none of these mechanisms has yet been verified experimentally.
The Role of Water Temperature in the Oceans
As we have seen, biodiversity in marine environments is greater in the tropics than it is at higher latitudes. However, this pattern does not appear to be driven by greater productivity in the tropics. Though marine productivity is greatest in temperate latitudes (see Figure 20.6), the high productivity in those regions is seasonal. As you may recall, temperature differences and stratification of the water in temperate zones make nutrients readily available during the seasonal mixing of the water column but scarce during periods when the water is stratified. In contrast, tropical marine environments experience relatively stable temperatures that lead to relatively small nutrient fluctuations and low but steady productivity. In 2010 researchers tested whether the patterns in species diversity across the oceans were better explained by productivity, mean water temperature, or variation in water temperature. They discovered that the only significant predictor of marine biodiversity across latitudes was the mean temperature of the sea surface. Because a higher mean temperature is a measure of greater total energy, this pattern further supports the energy–diversity hypothesis.
534
The patterns of species richness across and within latitudes are potentially affected by all three processes of habitat heterogeneity, temperature and precipitation on land, and mean temperature in the oceans. In all three cases, the mechanisms involved suggest that the global distribution of species richness is the outcome of an equilibrium between the processes that create new species and the processes that cause the extinction of existing species.